Research Report
Aerials and Roots Vegetative Tissues Differ in Guaiacol Peroxidases Responses to Cold Stress in Two Contrasting Medicago ciliaris Populations 
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2016, Vol. 7, No. 17 doi: 10.5376/mpb.2016.07.0017
Received: 17 Feb., 2016 Accepted: 02 Apr., 2016 Published: 19 Apr., 2016
Nourredine Y., Asma B., Chahrazed C., and Fatima Z.F.L., 2016, Aerials and Roots Vegetative Tissues Differ in Guaiacol Peroxidases Responses to Cold Stress in Two Contrasting Medicago ciliaris Populations,Molecular Plant Breeding, 7(17): 1-7(doi: 10.5376/mpb.2016.07.0017)
Effect of cold on antioxidant responses in aerial (shoots and leaves) and roots tissues of two contrasting accessions of M. ciliaris Krockers (Cil 126, tolerant and Cil 123, sensible) were investigated. Ten-day-old grown seedlings were subjected at different periods of cold acclimation (4°C) 2, 4, and 6 days. Peroxidases (POD) (EC 1.11.1.7) activities and isoenzymes expression of plantlets treated and control (23°C) of the aerial (shoot+leaves) and root vegetative tissues were carried. Global peroxidase activity under low temperature stress was higher in tolerant ecotype than sensible one. The same trend was also observed at expressed isoenzymes. On the other hand, whether in tolerant or sensitive, activity and isoenzymes peroxidase was more pronounced at the roots than in aerials tissues system.
Introduction
Cold stress is one of the major factors limiting agricultural productivity, consequently the plants evolved a numerous adaptive responses to cope with environmental stresses. Agricultural yield losses due to abiotic environmental stresses are obvious and have been well-documented (Cramer et al., 2011). Abiotic and biotic stress often causes a series of morphological, physiological, biochemical and molecular changes that unfavourably affect plant growth, development and productivity. Drought, salinity, extreme temperature (cold and heat) and oxidative stress are often interrelated; these conditions singularly or in combination induce cellular damage (Arun et al., 2011; Prince et al., 2013; Morina et al., 2015). Ghosh and Xu (2014) reviewed abiotic stress responses in plants and reported that these responses occur in various organ especially at roots tissues. Under normal growth condition, root absorbs water and nutrients from the soil and supplies them throughout the plant body, ensuring a cellular homeostasis balance. However, this cellular homeostasis altered during the stress period when roots tissues are forced to adopt several structural and functional modifications. Under cold stress a significant amount of reactive oxygen species (ROS) is produced, such as hydrogen peroxide (H2O2) and other dangerous derivatives oxygen, which cause progressive oxidative damage and as consequence cell death (Mohammadian et al., 2012). During cold acclimation, increases of enzymatic changes were associated with cold tolerance by a significant capture of these harmful molecules. It is often assumed that in many species, plants actively produce ROS as signaling molecules to control processes such as programmed cell death (Parent et al., 2008), abiotic stress responses, for example, in Barley (Baek et al., 2000), in rice (Kim and Thomas, 2011), in maize (Zeamays L.) (Lukatkin, 2002), pathogen defense, in Pepper (Wang et al., 2013). Higher plants contain numerous enzymatic and non-enzymatic reactive oxygen intermediate scavengers and antioxidants, both water and lipid soluble, localized in different cellular compartments (Mittler, 2002; Thakur and Nayyar, 2013). Antioxidant enzyme activities in plants are accepted as a good indicator of tolerance under stress conditions (Öklen et al., 2008).
Lee et al., (2009) argument that roots proteins increases highly under cold stress than in the leaves in rice. On the other hand, Cavalcanti et al., (2007) investigating the effect of salt stress in cowpea (Vigna unguiculata) plants, concluded that roots and leaves display distinct mechanisms of response during salt stress and recovery.
Radi´c et al., (2006) analyzing the peroxidase activity in leaves and roots under salt stress, highlighted that POD activity of salt-stressed plants changed with respect to different saline treatments and plant organs and this activity was mainly due to the de novo synthesis of the enzyme in shoot and root.
Lee and Lee (2000) analyzing the changes of antioxidant enzyme isoforms against chilling stress in the leaves of cucumber (Cucumis sativus L.) found five APX isoforms were presented in the leaves of cucumber. Furthermore, these authors showed that intensities of APX-4 and APX-5 were enhanced by chilling stress, whereas that of APX-3 was significantly increased in the post stress periods after chilling stress. Wang et al., (2009) suggested that peroxidase (POD) activity in normal conditions in two alfalfa cultivars was higher in root than in shoots. However, POD activities in shoot and root tissues of both cultivars showed similar level under chilling stress.
Keshavkant and Naithani (2001) reported that aerial parts of the chilling sensitive young sal (Shorea robusta) seedlings showed overproduction of reactive oxygen species. To gain in a clear understanding of the reactions exhibited by annuals Medicago plants under different duration of cold stress; we conducted a study with enzyme activity (peroxidase) and peroxidase isoenzymatique pattern as biochemical markers in response to cold stress in the aerial and roots tissues. These results may help in the development of a new variety of stress tolerant Medicago plants.
1 Results and Discussion
The biochemical changes observed in our study during cold stress in both vegetative tissues has been demonstrated in many species and under many stresses. Sánchez-Calderón et al., (2013) showed that in root vegetative system has to growth in media where the biotic and abiotic components are distributed heterogeneous. As soon as the root makes contact with the soil must sense and integrate biotic and abiotic cues in order to adjust their genetic program of post-embryonic root development. Ghosh and Xu (2014) reported that proteomic analyses of plant roots under various abiotic stress conditions revealed important information on proteins involved in the abiotic stress response. This leads to the identification of molecular and cellular mechanisms that are specific to certain abiotic stress or shared between two or more abiotic stress conditions. On the other hand, these authors highlighted that higher abundance of ROS scavengers was detected in roots under drought, high salinity and cold stress and can be looked upon as a preventive measure against oxidative damage caused due to high ROS levels.
To gain a better knowledge of the chilling response, Lee et al., (2009) suggested that it is imperative to analyze the tissue-specific proteome patterns under chilling stress. Prasad (1996) investigated the mechanisms involved in chilling injury or tolerance in developing maize seedlings subjected to low temperature, noted that peroxidase activities in acclimated plants increased gradually with 14 to 120 % with age and remained slightly higher during recovery in 2- to 4- days- old seedlings. Cavalcanti et al., (2007) comparing some antioxidative response during short-term salt stress, spotlighted that roots and leaves display distinct mechanisms of response during salt stress and recovery. However, these responses and/or the oxidative damage caused by oxygen species were not related with the growth reduction.
1.1 Changes in the activity of POD
Chilling tolerance or sensitivity in plants is well correlated with inherent antioxidant responses. Tolerant plant species generally have a better capacity to protect themselves from chilling-induced oxidative stress, via the enhancement of antioxidant enzyme activity. In plants, elevated ROS without an efficient scavenging system may lead to high injury index and finally death of the whole plant (Azzeme et al., 2016). These authors noted that in response to different levels of drought in the root and leaf tissues in oil palm plants the guaiacol POD activity in leaves was found to increase sharply under mild stress but dropped under moderate and further reduced under severe drought conditions even though the levels were still higher than that for the control. In the roots, the guaiacol POD activity significantly increased under mild and further increased moderate and severe stress conditions. Liu et al., (2013) have observed an enhancement in activities peroxidase synthesis pathway during cold acclimation. Yadegari et al., (2007) showed in soybean a significant changes in guaiacol peroxidase activity between cold acclimated and non-acclimates leaves 24 h after acclimation.
|
Figure 1A Aerial vegetative tissues peroxidase activity in M. ciliaris Krockers.ecotypes (Cil 126 and Cil 123) seedlings untreated (controls): (T02, T04, and T06) and treated with different durations (T2, T4, and T6) under cold stress. Data are shown as mean ± SD of three independent measurements |
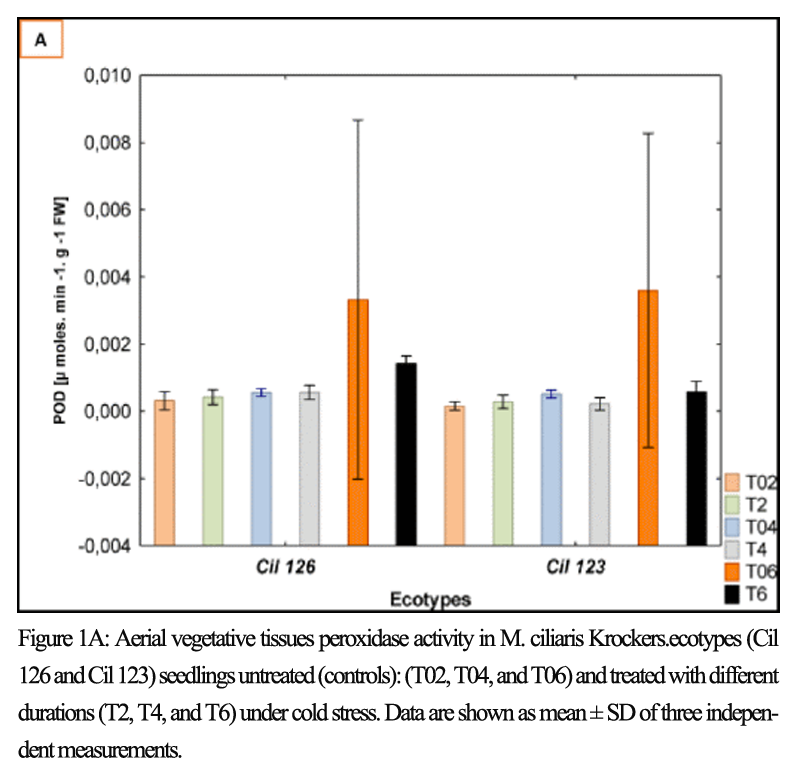
|
Figure 1B Root vegetative tissues peroxidase activity in M. ciliaris Krockers.ecotypes (Cil 126 and Cil 123) seedlings untreated (controls): (T02, T04, and T06) and treated with different durations (T2, T4, and T6) under cold stress. Data are shown as mean ± SD of three independent measurements |
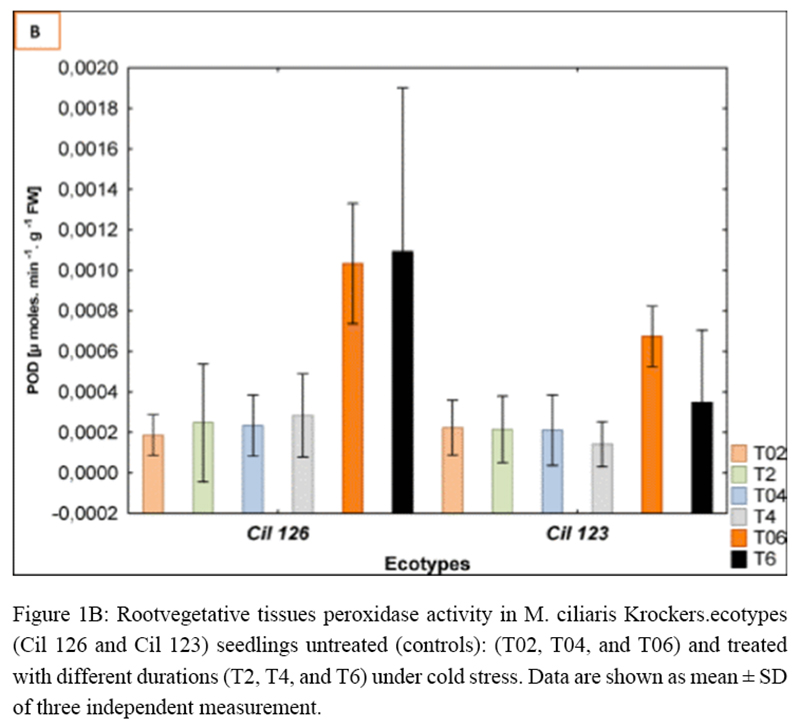
1.2 Effect of cold stress on peroxidase isoforms pattern
Guaicol peroxidases are known to utilize different substrates to metabolize H2O2. When guaiacol was used as a substrate, peroxidase activities were enhanced in chilling stressed-plant as compared with control plant (Lee and Lee, 2000). Comparing the peroxidase profiles between control plants and those treated under cold stress using native-page, the pattern of isoenzyme showed that cold treatment induced a high peroxidase activity. This activity increase slightly in roots tissues (Figure 2, A) than in aerial (leave and shoot) tissues (Figure 2, B) in both ecotypes. The peroxidase activities were more pronounced when cold stress is maintained in the time. This trend was observed in tolerant accession Cil 126 when plantlets were subjected at 6 days of stress. In contrast, in sensible accession Cil 123 peroxidase activities were slightly maintained (Figure 3: A and B). The same results were observed in chickpea (Cicer orientum L.) (Nazari et al., 2012). It was found that the number of bands (named POD–Rfs) was higher (five) in tolerant (Cil 126) (Figure 2) than in sensible (Cil 123) (Figure 3) with only four bands (POD 1 to POD 4). Moreover, results revealed that under cold treatment the intensity of bands were more pronounced in roots (Figure 2, A and Figure 3, A) than in aerial tissues (Figure 2, B and Figure 3, B). As showed in Figure 2A the intensity of POD 5-Rf_0.89 of Cil 126 and in Figure 3A POD 4-Rf_0.69 of Cil 123 in root was increased in cold treated plants in comparison of aerial tissues in both tolerant and sensible accession.
|
Figure 2 Peroxidase isoenzyme patterns in M. ciliaris Krockers. (Cil 126) seedlings untreated (controls): (T02, T04, and T06) and treated with different durations (T2, T4, and T6) under cold stress. |
.png)
|
Figure 3 Peroxidase isoenzyme patterns in M. ciliaris Krockers. (Cil 123) seedlings untreated (controls): (T02, T04, and T06) and treated with different durations (T2, T4, and T6) under cold stress |
.png)
Mohammadian et al., (2012) showed that in tissues of three cultivars of lemon (Citrusunshiu, Citrussinensis and Citrus lemon) the intensity of peroxidise bands is low in the sensible cultivars and is more important with 3 intense bands in both tolerant cultivars (lemon Citrussinensis and Citrus lemon) under a cold treatment. Furthermore, Lee and Lee (2000) have observed this tendency. These authors, analyzing the changes of antioxidant enzyme isoforms against chilling stress in the leaves of cucumber (Cucumis sativus L.), found five APX isoforms were presented in the leaves of cucumber. Furthermore, these authors showed that intensities of APX-4 and -5 were enhanced by chilling stress, whereas that of APX-3 was significantly increased in the post stress periods after chilling stress. On the other hand, Azzeme et al., (2016) noted that POD activity increased compared to control under different severity of drought in both tissues (leaves and roots) which showed that POD is an essential enzyme in scavening H2O2 in the oil palm seedlings. Mirzaee et al., (2013) reported an initial increase and later decrease in antioxidant activities in leaves and roots tissues of Brassica napus L. compared to control plants with the increasing duration of stress indicating the unbalance of the active oxygen metabolism system. The high accumulation of active oxygen was able to initiate and accelerate lipid peroxidation. Nevertheless, when the levels of the active oxygen exceeded the ability of antioxidant system to cope with them, damage to cellular components occurred (Cao et al., 2011).
2 Conclusion
Based on obtained results, it could be concluded that activities of peroxidase had different change trends with tolerant and sensible accession. The enhanced of scavenging ability for H2O2 in tolerant was better than in sensible accession. Two accessions Cil 126 and Cil 123 has potentially better antioxidant potential in roots tissues than in aerial tissues. Cold adversely affects the plant metabolism in Medicago populations seedlings by affecting antioxidant enzyme activities in leaves and roots tissues. Thus, this work can be benefited as potential targets for enhancement of stress tolerance in Medicago species under different durations, through genetic of mRNAs transcripts engineering and breeding programmes to develop cold tolerance local annuals Medicago cultivars.
3 Materials and Methods
The study was carried on two annual Medicago ciliaris Krockers (Cil 126, tolerant and Cil 123, sensible) (Table 1). Ten seeds for each accession were germinated after scarification and disinfected by dipping in 70% (v/v) ethanol, at temperature room in Petri dishes containing universal compost imbibed with distiller water. At three days growth stage, seedlings were divided into two lots. Cold treated lot at 4°C for three durations 2, 4 and 6 days (T2, T4 and T6) and control lot kept at 23°C (T02, T04 and T06). The experimental layout was a completely randomized block design with 3 replications.
|
Table 1 Accessions analyzed for peroxidase antioxidant under cold stress with their origin and ecological description |

Aerial (shoot and leaves) and root tissues from control and treated plants (Figure 4) were homogenized in homogenization buffer (10 mM Tris-KCl, pH 6.8, 10% (w/v) saccharose and 1 mM PMSF).Tissues were frozen in liquid nitrogen and ground in mortar on ice using homogenization buffer.
|
Figure 4 Aerial (shoot and leaves) and root tissues from control and treated plantlets |

Peroxidase activity (EC 1.11.1.7) (POD) was assayed in reaction solution (3 ml) containing 0.2 M sodium acetate buffer pH 4.6, 1% o-dianizidine, 10% H2O2 (Mac Adam et al., 1992) modified. The reaction was started by adding 10 µl crude extract, and the enzyme activity is monitored for every 15 seconds for 3 minutes using a spectrophotometer at 460 nm as o-dianizidine oxidation, with 3 replications. Enzyme activity was expressed as µmole of o-dianizidine oxidation. FM/ming of fresh matter (FM).
The aerial (shoot and leaves) and root tissues peroxidase pattern were resolved by native in 10% separating polyacrylamide gelelectrophoresis on vertical slab gel (Hoffer, USA) using the procedure of Laemmli (1970). Equal amounts of proteins (15 µL) were loaded on to each lane. The gel electrophoresis experiment was repeated tree times. To determine the pattern of peroxidase isoforms gels were staining and visualized by immersing the gels in 100 mL 0.2 M acetate buffer pH 4.6 added with 1% o-dianizidine (Sigma), dissolved in 2 mL 95% ethanol, and 200 µL 3% H2O2 at room temperature until the brown color appeared. Scanned Isozymes profiles gels were analyzed by Software GELANALYZER (Istvan Lazar Hungary Copyright 2010) to determine Rfs bands.
Acknowledgments
This work was supported by Research PNR projectentitled ″Biotechnologie des Rhizobiums et Amélioration des Plantes″ (code Project 61) affiliated to the Algerian Ministry of Higher Education and Scientific Research. We are very grateful two anonymous referees and editor of this paper for their helpful and constructive comments.
Authors' contributions
YN conceived of the study, design it has and performed the statistical analysis and analysis results. BA and CC carried out the research as part of their MSc project. F-LFZ have been involved in the revising it critically for intellectual content. All authors have participated sufficiently in the work and all authors read and approved the final manuscript.
Arun K.S. and Venkateswarlu B., 2011, Abiotic stress response in plants-Physiological, Biochemical and Genetic Perspectives, Edited by Arun Kumar Shanker and B. Venkateswarlu, pp.346, Published by InTechJaneza Trdine 9, 51000 Rijeka, Croatia
Azzeme A.M., Abdullah S.N.A., Aziz M.A., and Wahab P.E.M., 2016, Oil palm leaves and roots differ in physiological response, antioxidant enzyme activities and expression of stress-responsive genes upon exposure to drought stress, Acta Physiol. Plant, 38(2): 1-2
http://dx.doi.org/10.1007/s11738-016-2073-2
Baek S-H., Kwon I-S., Park T-I., Yun S.J., Kim J.K. and Choi K.-G., 2000, Activities and isozyme profile of antioxidant enzymes intracellular compartment of overwintering Barley leaves, Journal of Biochemistry and Molecular Biology, 33 (5): 385-390
Cao H., Sun C., Shao H. and Lei X., 2011, Effects of low temperature and drought on the physiological and growth changes in oil palm seedlings, African Journal of Biotechnology, 10(4): 2630–2637
Cavalcanti F.R., Lima J.P.M.S., Ferriera-Silva S.L., Viégas R.A. and Silveira J.A.G., 2007, Roots and leaves display contrasting oxidative response during salt stress and recovery in cowpea, Journal of Plant Physiology 164(5): 591-600
http://dx.doi.org/10.1016/j.jplph.2006.03.004
Cramer G.R., Urano K., Delrot S., Pezzotti M. and Shinozaki K., 2011, Effects of abiotic stress on plants: a systems biology perspective, BMC Plant Biology, 11(1): 1-14
http://dx.doi.org/10.1186/1471-2229-11-163
Gaspar Th., Kevers C., Hausman J.F., Berthon J. and Rippeti V., 1992, Practical uses of peroxidase activity as predictive marker of rooting performance of micro propagated shoots, Agronomie,12(10): 757-765
http://dx.doi.org/10.2298/ABS0802223M
Ghosh D. and Xu J., 2014, Abiotic stress responses in plant roots: a proteomics perspective, Frontiers in Plant Science, 5(6): 1-13
http://dx.doi.org/10.3389/fpls.2014.00006
Keshavkant S. and Naithani S.C., 2001, Chilling-induced oxidative stress in young sal (Shorea robusta) seedlings. Acta Physiologiae Plantarum, 23(4): 457-466
http://dx.doi.org/10.1007/s11738-001-0056-3
Kim S.-I. and Thomas H.T., 2011, Evaluation of seedling cold tolerant in rice cultivars: a comparison of visual ratings and quantitative indictors of physiological changes, Euphytica, 178(3): 437-447
http://dx.doi.org/10.1007/s10681-010-0343-4
Kuk, Y.I.,Shin J.S.,Burgos N.R., Hwang T.E., Han O., Cho B.H., Jung S. and Guh J.O., 2003, Antioxidative Enzymes Offer Protection from Chilling Damage in Rice Plants, Crop Science Society of America 43(6): 2109-2117
http://dx.doi.org/10.2135/cropsci2003.2109
Kukavica B., Mitrović A., Mojović M. and Veljović-Jovanović S., 2007, Effect of indole-3-acetic acid on pea root growth, peroxidase profiles and hydroxyl radical formation. Arch. Biol. Sci. (Belgrade) 59(4): 319-326
http://dx.doi.org/10.2298/ABS0704319K
Laemmli U.K., 1970, Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature, 227(5259): 680-685
http://dx.doi.org/10.1038/227680a0
Larkindale J., Hall J.D., Knight M.R. and Vierling E., 2005, Heat stress phenotypes of Arabidopsis mutants implicate multiple signalling pathways in the acquisition of thermotolerance, Plant Physiology, 138(2): 882–897
http://dx.doi.org/10.1104/pp.105.062257
Lee D.-G., Ahsan N., Lee S.-H., Lee J.J., Bahk J.D., Kang K.Y. and Lee B.-H., 2009, Chilling stress-induced proteomic changes in rice roots, Journal of Plant Physiology, 166(1): 1-11
http://dx.doi.org/10.1016/j.jplph.2008.02.001
Lee H.D., and Lee C.B., 2000, chilling stress-induced changes of antioxidant enzymes in the leaves of cucumber: in gel enzyme activity assays, Plant Science 159 (1): 75-85
http://dx.doi.org/10.1016/S0168-9452(00)00326-5
Liu W., Yu K., He T., Li F., Zhang D. and Liu J., 2013, The Low Temperature Induced Physiological Responses of Avena nuda L., a Cold-Tolerant Plant Species, The Scientific World Journal, 2013(2013):658793-658793
http://dx.doi.org/10.1155/2013/658793
Lukatkin A.S., 2002, Contribution of oxidative stress to the development of cold-induced damage to leaves of chilling-sensitive plants: 1. Reactive oxygen species formation during plant chilling, Russian Journal of Plant Physiology, 49(49):622-627
http://dx.doi.org/10.1023/A:1020232700648
Mac Adam J.W., Nelson C.J., and Sharpe R.E., 1992, Peroxidase activity in the leaf elongation zone of tall fescue, Plant Physiology. 99: 872-878
http://dx.doi.org/10.1104/pp.99.3.872
Mittler R., 2002, Oxidative stress, antioxidants and stress tolerance, Trends in Plant Science, 7(9):405–410
http://dx.doi.org/10.1016/S1360-1385(02)02312-9
Mirzaee M., Moieni A. and Ghanati F., 2013, Effects of drought stress on the lipid peroxidation and antioxidant enzyme activities in two canola (Brassicanapus L.) cultivars, Journal of Agricultural Science & Technology, 15(3):593-602
Mohammadian M.A., Largani Z.K. and Sajedi R.H., 2012, Quantitative and qualitative comparison of antioxidant activity in the flavedo tissue of three cultivars of citrus under cold stress, Australian Journal of Crop Science, 6(3): 402-406.
Morina F., Vidovic M., Kukavica B. and Veljovic-Jovanovic S., 2015, Induction of peroxidase isoforms in the roots of two Versbascumthapsus L. populations involved in adaptive responses to excess ZN2+ and Cu2+, Botanica Serbica, 39(2): 151-158
Nazari M., Maali Amiri R., Mehraban F.Z. and KhaneghahH.Z., 2012, Change in antioxidant responses against oxidative damage in black chickpea following cold acclimation. Russian Journal of Plant Physiology. 59 (2): 183-189
http://dx.doi.org/10.1134/S102144371201013X
Öklen H.A., Eyidoðan F., Demirba D., Bayraç A.T., Öz M.T., Özgür E., Selçuk F. and Yücel M., 2008, Antioxidant Responses of Lentil to Cold and Drought Stress, Plant Biochemistry & Biotechnology, 17(1): 15-21
http://dx.doi.org/10.1007/BF03263254
Okuda T., Matsuda Y., Yamanaka A. and Sagisaka S., 1991, Abrupt increase in the level of hydrogen-peroxide in leaves of winter wheat is caused by cold treatment, Plant Physiology, 97(3):1265-1267
http://dx.doi.org/10.1104/pp.97.3.1265
Prasad T.K., 1996, Mechanisms of chilling-induced oxidative stress injury and tolerance in developing maize seedlings: changes in antioxidant system, oxidation of proteins and lipids, and protease activities, The Plant Journal, 10(6): 1017-1026
http://dx.doi.org/10.1046/j.1365-313X.1996.10061017.x
Prince S.J., Mutava R.N. and Nguyen H.T., eds., C. Kole., 2013 Genomics and Breeding for Climate-Resilient Crops, Root characters, pp.67-131
Parent C., Capelli N. and Dat J., 2008, Formes reactive de l’oxygène, stress et mort cellulaire chez les plantes, Comptes Rendus Biologies, 331(4): 255-261
http://dx.doi.org/10.1016/j.crvi.2008.02.001
Radi´c S., Radic´-Stojkovic M. and Pevalek-Kozlina B., 2006, Influence of NaCl and manitol on peroxidase activity and lipid peroxidation in Centaurea ragusina L. roots and shoots, Journal of Plant Physiology, 163(12): 1284-1292
http://dx.doi.org/10.1016/j.jplph.2005.08.019 PMid:17126732
Sánchez-Calderón L., Ibarra-Cortés M.E. and Zepeda-Jazo I., 2013, Root Development and Abiotic Stress Adaptation, Abiotic Stress - Plant Responses and Applications in Agriculture, pp.135-168
http://dx.doi.org/10.5772/55043
Thakur P. and Nayyar H., eds., Narendra T., and Sarvajeet S.G., 2013, Facing the Cold Stress by Plants in the Changing Environment: Sensing, Signalling, and Defending Mechanisms, Plant Acclimation to Environmental Stress, pp.29-69
Wang W.B., Kim Y.H., Lee H.S., Deng X.P. and Kwak S.S., 2009, Differential antioxidation activities in two alfalfa cultivars under chilling stress, Plant Biotechnology Reports, 3(4): 301-307
http://dx.doi.org/10.1007/s11816-009-0102-y
Wang J.E., Liu K.K., Li D.W., Zhang Y.L., Zhao Q., He Y.M. and Gong Z.H., 2013, A novel peroxidase Can-POD gene of Pepper is involved in defense responses to Phytophtoracapsici infection as well as abiotic stress tolerance, International Journal of Molecular Sciences, 14(2): 3158-3177
http://dx.doi.org/10.3390/ijms14023158 PMid:23380961 PMCid:PMC3588037
Yadegari L.Z., Heidari R. and Carapetian J., 2007, Cold pre-treatment-Induced Changes in antioxidant Enzyme Activities and relative Water Content and Soluble Sugars in shoots and Roots of Soybean Seedlings, Journal of Biological Sciences, 7(8): 1525-1530



. PDF(350KB)
. FPDF
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Nourredine Yahia
. Asma Bouira
. Chahrazed Cheriet
. Fatima Zohra Fyad-Lameche
Related articles
. Peroxidase activity
. Isoenzymes
. Cold stress
. M. ciliaris
. PAGE electrophoresis
Tools
. Email to a friend
. Post a comment