Research Article
Variability, Heritability and Genetic Advance Analysis for Some Morphological Characters in Oilseed Brassica Breeding Lines 
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2016, Vol. 7, No. 20 doi: 10.5376/mpb.2016.07.0020
Received: 05 Apr., 2016 Accepted: 08 May, 2016 Published: 17 May, 2016
Yared S., and Misteru T., 2016, Variability, Heritability and Genetic Advance Analysis for Some Morphological Characters in Oilseed Brassica Breeding Lines, Molecular Plant Breeding, 7(20): 1-8 (doi: 10.5376/mpb.2016.07.0020)
Sixty four Brassica breeding lines were investigated for some morphological characters to identify the extent and nature of genetic variability and determine the heritability and genetic advance during 2014 cropping season. Analysis of variance showed the existence of considerable genetic variation among the lines for further selection and hybridization efforts. The maximum number of secondary branches/plant was observed by the breeding line code≠64. The highest yield/plot was recorded by the breeding line code≠48 followed by the breeding line code≠25 and code≠64. Breeding line code≠53 exhibited the maximum 1 000 seed weight. Cluster analysis categorized the breeding lines into nine clusters. All lines were grouped regardless of their pedigree record which might be the result of selection pressure applied on different morphological characters. 71.1% of the variation was explained by the first three principal components as described by the principal component analysis. Number of secondary branches/plant and yield/plot were among the major positive contributor while 1 000 seed weight had negative contribution in the first principal component in which 32% of the variation was explained. These characters also recorded high heritability values in broad sense along with high genetic advance as percent of mean for which early generation selection would be effective in improvement program.
Introduction
Brassica carinata commonly called Ethiopian mustard predominantly cultivated in Ethiopia since ancient time, which is used for oil, condiments, medicines and vegetables (Riley and Belayneh, 1982). It covered 44 041 hectares of land, and produced 62 450.27 tons with productivity of 1.42 ton/ha (CSA, 2013/14). Brassica napus was also grown in state farms in 1990s, but it could not be sustained due to diseases outbreak such as blackleg (Leptosphaeria maculans). Currently, some hybrid and open pollinated varieties of B. napus and B. juncea, which are canola type and disease resistant, were introduced by private companies and tested for adaptation, and as a result, some varieties have been registered by National Variety Release Committee (NVRC) for production in the country. However, such varieties are very expensive to procure and costly to be grown by small scale farmers in developing countries like Ethiopia. Despite unsuccessful efforts in developing canola type B. carinata so far, in order to tap the potential of this crop as an oilseed, developing disease resistant, canola type and high yielding varieties with desirable agronomic traits remains the primary objective of oilseed Brassica breeding in Ethiopia. As genetic diversity is a prerequisite for any breeding program of developing improved varieties or hybrids, understanding the extent and nature of this diversity is essential for enhancing the breeding efforts such as selection and hybridization. Several researches on the extent and nature of genetic variation have been done for various crops including Brassica species (Mailer et al., 1997; Alemayehu and Becker, 2002; Mahasi and Kamundia, 2007; Abbas et al., 2008; Uysal et al., 2010; Golkar et al., 2011; Khodadadi et al., 2011; Žiarovská et al., 2012; Ahmad et al., 2013; Kumar et al., 2013; Nasim et al., 2013; Iqbal et al., 2014; Shafari et al., 2015). Heritability and genetic advance studies were also done by several researchers (Ali et al., 2013; Nasim et al., 2013; Bind et al., 2014; Synrem et al., 2014). The objective of this study was to identify the extent and nature of genetic variability and determine the heritability and genetic advance for morphological characters of oilseed Brassica breeding lines.
1 Results
Analysis of variance shows that breeding lines were significantly different (p<0.05 and p<0.01) for all characters under study (Table 1). Variability among the breeding lines for the characters measured was also estimated using Duncan Multiple Range Test (DMRT) (Table 2). Breeding line code 20 was early to flower and mature (48 and 144 days, respectively). Breeding line code 23 (code≠hereafter) had the maximum height (231 cm) while code≠50 recorded the least height (149.5 cm). The maximum number of primary branches/plant (11.2) was recorded by the code≠27, where as the maximum number of secondary branches/plant (18.8) was obtained by the code≠64. Code≠64 had also recorded the highest number (248) of pods/plant. The maximum yield/plot was recorded by code≠48 (1 291 kg/ha) followed by code≠25 (1 277.4 kg/ha) and code≠64 (1 102.1 kg/ha). The highest 1 000 seed weight was recorded by the code≠53.
|
Table 1 Mean squares of the analysis of variance for morphological characters in oil seed Brassica breeding lines |
.png)
|
Table 2 Mean performance of the breeding lines for morphological characters under study |

1.1 Cluster analysis
Clustering indicates that all breeding lines were grouped into nine clusters. Solitary line was found in the first, second, third, and fifth cluster, where as fourth, sixth, seventh, eighth, and ninth cluster consisted of 4, 3, 13, 20, and 20 breeding lines, respectively (Table 3 and Figure 1).
|
Table 3 Grouping of oilseed Brassica breeding lines in different clusters |
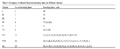
|
Figure 1 Average minimum variance dendrogram which shows the numbers of breeding lines clustered based on morphological characters studied |
.png)
1.2 Principal component analysis
Principal component analysis indicates that 71.1% of the variation was explained by the first three principal components (Table 4). Characters such as days to flowering, days to maturity, number of secondary branches/plant, number of pods/plant, and yield/plot showed the major positive contribution under the first principal component in which about 32% of the variation explained, where as 1 000 seed weight showed negative weight under this principal component. Plant height, number of primary branches/plant, and 1 000 seed weight had the major positive contribution in the second principal component, whereas number of secondary branches/plant and number of pods/plant recorded negative weight under this component by which 26.4% of the variation was explained. In principal component III (12.8% of variation), plant height, number of primary branches/plant, number of secondary branches/plant, number of pods/plant, and 1 000 seed weight had positive weight while days to flowering, days to maturity, and yield/plot showed negative weight.
|
Table 4 Component scores of the first three principal components of the breeding lines |

1.3 Genetic parameters estimate
High phenotypic and genotypic variances was recorded for yield/plot, number of pods/plant, and plant height (Table 5). Maximum phenotypic and genotypic coefficients of variation were recorded by number of secondary branches/plant (0.680 and 0.625, respectively) followed by yield per plot (0.425 and 0.389, respectively) and 1 000 seed weight (0.232 and 0.225, respectively). Heritability in the broad sense estimates were grouped into high (>50%), moderate (20-50%), and low (<20%) as described by Stansfield (1988). High heritability value in broad sense was observed for all characters except days to flowering which showed heritability value of 40.62%. High heritability along with high genetic advance as percent of mean was recorded by number of secondary branches/plant followed by yield/plot and number of pods/plant
|
Table 5 Component of variance, coefficient of variation, heritability in broad sense and genetic advance for morphological characters studied |

2 Discussion
Variance analysis shows the existence of genetic variability among the lines from which selection can be done to develop hybrid varieties with desirable agronomic characters. Similarly, Kumar et al. (2013) studied genetic diversity for agro-morphological and oil quality traits in 46 genotypes of Indian mustard, and showed the presence of considerable variability among the genotypes. Likewise, Synrem et al. (2014) studied the genetic variability in intra specific crosses of Indian mustard Brassica juncea (L.) Czern and Coss. Genotype, indicated that significant differences among the crosses were found for all the characters under study except length of siliqua. Significant variation for plant height, number of primary branches/plant, number of secondary branches/plant, number of pods/plant, and seed yield per plot was also reported (Khan et al., 2006).
All breeding lines were grouped regardless of their pedigree. This indicates that diversity were present within breeding lines of the same pedigree while similarity was found among the lines with the different pedigree. This might be revealed as a result of the differences in selection pressure exerted on different characters. Genetic diversity among lines indicates differences in gene frequencies which may result in heterotic group. Khodadadi et al. (2011) studied 36 winter wheat genotypes, and categorized them into seven groups.
Principal component analysis revealed the pattern of genetic variation existed. According to this study, the majority of variation was explained by the first three principal components. The principal component analysis in Khodadadi et al. (2011) indicated that the first five components explained over 97% of the variation in 36 wheat genotypes.
Values of phenotypic variance were greater than genotypic variance for all characters under study which is similar to the result in Kumar et al. (2013). Synrem et al. (2014) reported maximum genotypic and phenotypic coefficients of variation for number of secondary branches/plant, seed yield/plant, and number of primary branches/plant. High value of heritability for plant height, number of primary branches/plant, number of secondary branches/plant, number of pods/plant, and seed yield/plot was also recorded in Brassica napus (Khan et al., 2006 ). In Brassica juncea, high heritability estimates in broad sense were recorded for 1 000 seed weight, days to maturity, days to flowering, plant height, and main shoot length (Bind et al., 2014). Iqbal et al. (2014) reported high heritability value for plant height, number of primary branches/plant, number of secondary branches/plant, number of siliqua/plant, 1 000 seed weight, and days to maturity. The present study showed that yield/plot, number of pods/plant, and plant height exhibited high value of genetic advance. This indicates that these characters involved additive gene action in their expressions, and can be improved through early generation selection. High value of heritability coupled with high genetic advance as percent of mean was observed for number of secondary branches/plant, seed yield/plant, and number of primary branches/plant in Brassica juncea (Synrem et al., 2014).
3 Conclusion
This study has revealed the existence of considerable genetic variation among the breeding lines for the characters under study, which can be used for further selection and hybridization. High heritability estimates in broad sense along with high genetic advance as percent of mean was observed for number of secondary branches/plant, yield/plot, and 1 000 seed weight. As additive gene action is pronounced in the expression of these characters, early generation selection would be effective in breeding program.
4 Materials and Methods
This experiment was carried out at Holetta Agricultural Research Center during the growing season of 2014. Sixty four breeding lines (S6) were grown in 8x8 simple lattice designs with two replications. These selfed lines were derived from various crosses and cultivars of oilseed Brassica (Table 6). Oilseed Brassica lines other than Ethiopian mustard (Brassica carinata) lines were introduced from abroad for breeding purpose. Each line was planted in a plot consisting of 6 rows of 3 m length and 30 cm between rows. Data were recorded on ten randomly selected plants from central 4 rows for plant height, number of primary branches/plant, number of secondary branches/plant, number of pods/plant and 1 000 seed weight. Days to flowering, days to maturity and seed yield were recorded on plot basis. All the recommended agronomic practices were followed for good establishment of the crop. The relative efficiency of simple lattice design over randomized complete block design (RCBD) was less than 25%, therefore, analysis of variance (Table 1) for the characters under study considered the RCBD model as suggested by Bolanŏs and Edmeades (1996). Analysis of variance, mean performance, cluster, and principal component were done using SAS software version 9.00 (SAS, 2002). Phenotypic and genotypic coefficients of variation were calculated following Burton and De Vane (1953). Broad sense heritability of the characters was computed as suggested by Allard (1960), and the expected genetic advance under selection assuming selection intensity of 5% (2.063) was determined following Johnson et al. (1955).
|
Table 6 Sources of breeding lines of oilseed Brassica |
Authors’ contribution
The first author has done the analysis and interpretation of data as well as the manuscript draft while the second author has made substantial contributions to conception, design and acquisition of data as well as revision of the final manuscript.
Acknowledgments
Authors are grateful for Highland Oil Crops Commodity Research technical staffs for their relentless effort in collecting data for this experiment.
Abbas S.J., Farhatullah, Khan I.A., Marwat K.B., and Munir I., 2008, Molecular and biochemical assessment of brassica napus and indigenous campestris species, Pakistan Journal of Botany, 40(6): 2461-2469
Ahmad B., Mohammad S., Azam F., Ali I., Ali J., and Rehman S., 2013, Studies of Genetic Variability, Heritability and Phenotypic Correlations of Some Qualitative Traits in Advance Mutant Lines of Winter Rapeseed (Brassica napus L.), American-Eurasian Journal of Agricultural & Environmental Science, 13(4): 531-538
Alemayehu N., and Becker H.C., 2002, Genotypic diversity and pattern of variation in a germplasm material of Ethiopian mustard (Brassica carinata A. Brun), Genetic Resource and Crop Evolution, 49(6): 573-582
http://dx.doi.org/10.1023/A:1021204412404
Ali Y., Farhatullah, Rahman H., Nasim A., Azam S.M., and Khan A., 2015, Heritability and correlation analysis for morphological and biochemical traits in Brassica carinata, Sarhad Journal of Agriculture, 29(3): 359-370
Allard R.W., eds., 1960, Principles of Plant Breeding, John Wiley & Sons, Inc., New York, pp. 75-88
Bind D., Singh D., and Dwivedi V.K., 2014, Genetic variability and character association in Indian mustard [Brassica juncea (l) czerns & coss], Agricultural Science Digest, 34(3): 183–188
http://dx.doi.org/10.5958/0976-0547.2014.00998.7
Bolaňos J., and Edmeades G.O., 1996, The importance of anthesis-silking interval in breeding for drought tolerance in tropical Maize, In: G.O. Edmeades, M. Bänziger, H.R. Mickelson and C.B, Peňa-Valdivia (eds.), Developing drought and low N-tolerant Maize, Proceedings of a symposium, March, 25-29, 1996, CIMMYT, El Batán, Mexico
http://dx.doi.org/10.1016/0378-4290(96)00036-6
Burton G.W., and De Vane E.H., 1953, Estimating heritability in all fescue (Festuca arundinacea) from replicated clonal material, Agronomy Journal, 45(10): 478-481
http://dx.doi.org/10.2134/agronj1953.00021962004500100005x
CSA, 2013/14, Agricultural sample survey: Report on area and production of major crops, Volume I, Central Statistical Agency (CSA) of the Federal Democratic Republic of Ethiopia, Addis Ababa, Ethiopia.
Golkar P., Arzani A., and Rezaei A.M., 2011, Genetic Variation in Safflower (Carthamus tinctorious L.) for Seed Quality-Related Traits and Inter-Simple Sequence Repeat (ISSR) Markers, International Journal of Molecular Science, 12(4): 2664-2677
http://dx.doi.org/10.3390/ijms12042664 PMid:21731465 PMCid:PMC3127141
Iqbal S., Haque S., Nath U.K., and Hamim I., 2014, Genetic diversity analysis of mustard germplasm based on phenotypic traits for selection of short duration genotypes, International Journal of Agricultural Science Research, 3(8): 141-156
Johnson H.W., Robinson H.F., and Comstock R.E., 1955, Estimates of genetic and environmental variability in soybeans, Agronomy Journal, 47(7): 314–318
http://dx.doi.org/10.2134/agronj1955.00021962004700070009x
Khan F.A., Ali S., Shakeel A., Saeed A., and Abbas G., 2006, Genetic variability and genetic advance analysis for some morphological traits in B. napus L., Journal of Agricultural Research, 44(2): 83-88
Khodadadi M., Fotokian M.H., and Miransari M., 2011, Genetic diversity of wheat (Triticum aestivum L.) genotypes based on cluster and principal component analyses for breeding strategies, Australian Journal of Crop Science, 5(1): 17-24
Kumar B., Pandey A., and Singh S.K., 2013, Genetic diversity for agro-morphological and oil quality traits in Indian mustard (Brassica juncea czern & coss), The bioscan, 8(3): 771-775
Kumar A., Banga S.S., Meena P.D., and Kumar P.R., eds., 2015, Brassica breeding: Breeding and management, CAB International, London, UK, pp.1-10
http://dx.doi.org/10.1079/9781780644837.0001
http://dx.doi.org/10.1079/9781780644837.0000
Mahasi M.J., and Kamundia J.W., 2007, Cluster analysis in rapeseed (Brassica napus L.), African Journal of Agricultural Research, 2(9): 409-411
Mailer R.J., Wratten N., and Vonarx M., 1997, Genetic diversity among Australian canola cultivars determined by random amplified polymorphic DNA, Australian Journal of Experimental Agriculture, 37(7): 793-800
http://dx.doi.org/10.1071/EA97059
Nasim A., farhatullah, Iqbal S., Shah S., and Azam S.M., 2013, Genetic variability and correlation studies for morphophysiological traits in Brassica napus, Pakistan Journal of Botany, 45(4): 1229-1234
Riley K.W., and Belayneh H., 1982, Report from an oil crop collection trip in Ethiopia, Cruciferae Newsletter, 7: 5–6
SAS Institute, 2002, SAS/STAT guide for personal computers, version 9.00 edition. Cary, NC: SAS Institute Inc.
Sharafi Y., Majidi M.M., Jafarzadeh M., and Mirlohi A., 2015, Multivariate analysis of genetic variation in winter rapeseed (Brassica napus L.) cultivars, Journal of Agricultural Science and Technology, 17(4): 1319-1331
Stansfield W.D., 1988, Theory and problems of genetics, M. C. Grow Hill Book Co. New York USA, pp.220-221
Synrem G. J., Rangare N.R., Myrthong I., and Bahadure D.M., 2014, Variability studies in Intra specific crosses of Indian mustard [Brassica juncea (L.) Czern and Coss.] genotypes, IOSR Journal of Agriculture and Veterinary Science, 7: 29-32
http://dx.doi.org/10.9790/2380-07932932
Uysal H., Fu Y.B., Kurt O., Peterson G.W., Diederichsen A., and Kusters P., 2010, Genetic diversity of cultivated flax (Linum usitatissimum L.) and its wild progenitor pale flax (Linum bienne Mill.) as revealed by ISSR markers, Genetic Resource and Crop Evolution, 57(7): 1109-1119
http://dx.doi.org/10.1007/s10722-010-9551-y
Žiarovská J., Ražná K., Senková S., Štefúnová V., and Bežo M., 2012, Variability of linum usitatissimum l. based on molecular markers, Journal of Agricultural and Biological Science, 7: 50-58



. PDF(324KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Yared Semahegn
. Misteru Tesfaye
Related articles
. Brassica breeding lines
. Analysis of variance
. Cluster analysis
. Principal component analysis
. Heritability
. Genetic advance as percent of mean
Tools
. Email to a friend
. Post a comment