Research Article
Wide Compatibility Gene Approaches and Heterosis Relationship in Japonica x Indica Hybrid Rice (Oryza sativa L.) 
2 Indian Institute of Agricultural Biotechnology, Garhkhatanga, Ranchi, Jharkhand, 834010, India
3 Division of Genomic Resources,National Bureau of Plant Genetic Resources, Pusa Campus, New Delhi, 110012, India
4 Division of Biotechnology, Indian Institute of Rice Research, Rajendranagar, Hyderabad, 500030 (Telangana), India
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2016, Vol. 7, No. 21 doi: 10.5376/mpb.2016.07.0021
Received: 27 Apr., 2016 Accepted: 03 May, 2016 Published: 01 Jun., 2016
A special class of rice germplasm, referred as wide-compatible can produce hybrids with enhanced spikelet fertility when crossed with japonica×indicia rice lines having neutral allele. Present study was conducted to identify the presence of neutral allele and relationship in the hybrid performance. The wide-compatible genotype screening resulted positive selection having neutral allele and responsible for increased spikelet fertility with possible utilization as donor parent in hybrid development programme for achieving higher yield. Combining ability and heterosis for grain yield and its component traits derived from japonica×indica experimental hybrids revealed higher general combining ability variance than their respective specific combining ability variances indicating the predominance of additive gene action, opened the way for fixing the yield contributing gene by breeding through transgressive selection. Most of the experimental hybrids showed significant and positive SCA effects for grain yield as well as several other yield related component traits. Wide compatibility gene investigation clearly demonstrated the utility of InDel primer in the identification of wide compatible genotypes possessing neutral allele at S5 locus. The genotypes having natural allele (281-bp) can give rise to fully fertile panicle and significant contribution towards yield and heterosis. Association index between genetic diversity and three SH of GY hybrids on the basis of microsatellite data set surprisingly exhibited significant and positive correlation indicated the possible utility of microsatellite markers for successful utilization and establishing the strong heterotic relationship.
Introduction
Rice (Oryza sativa L.) is life and known as prince among the all cereals. More than half of the world’s population depends on rice for calories and protein, especially among the developing countries 90 per cent of the rice is grown and consumed in Asia (rice bowl of the world), where 60 percent of the earth’s people and two third of world’s poor live (Khush and Virk, 2000). Moreover, drastic depletion of land and water resources due to population explosion and urbanization demands for a promising and economically viable technology to achieve higher productivity to meet the future food demand of 852 million tons in 2035. Ensuring the demand, we need to enhance rice production to current production level of 676 million tonns by increasing of 5 to 8 million tons production every year to fillip the dietary requirements of ever growing populations Khush (2013). Hybrid technology is one of the most feasible options to full-fill the demands of increasing population by developing new high yielding hybrids and improved lines/varieties with stable performance for different agro-climatic conditions. The most common, easy and effective means for developing and identifying new hybrids depends on strong wide combining ability and usefulness to assess nicking ability in crops and at the same time elucidate the nature and magnitude of gene actions involved, provides breeders about insight of nature and relative magnitude of fixable and non-fixable genetic variances ie. dominance or epistatic components. Wider combining ability provides useful information for the selection of donor parents for effective breeding programme. Such information are required to design efficient breeding strategy for rapid dynamic crop improvement. The alternate discovery of major wide compatible QTLs located on S5 locus of chromosome no. 6 for rapid detection of wide compatible genotypes and relationship between different agronomic QTLs with genomic diversity provided a powerful tool for assessing the hybrid performance and capable of distinguishing the widely compatible japonica and indica rice genotype for utilizing as donor for developing fully fertile hybrids and targeted introgression in to elite rice hybrids Sundaram et al. (2010).
1 Results and Analysis
1.1 Analysis of variance
ANOVA for combining ability (Table 1) revealed mean square due to lines were significant for all the fifteen characters under study. Variance due to line × tester fashion was highly significant except for PBT, FLA, HI and LBR (Saleem et al., 2010). The variance study suggested importance of both additive and non-additive gene effects represented by general and specific combining ability variances, for majority of the traits. The GCA variances were higher than the corresponding SCA variances for almost the traits indicated preponderance of additive gene effects for maintaining heterozygosity for improving desirable traits. Hence, the suitable breeding strategy for attaining high yield would be the full or partial exploitation of hybrid vigor through development of synthetic or composite cultivars. The estimates of average degree of dominance was greater to unity emphasized predominance of additive gene effects. The almost unity value of predictability ratio suggested prevalence of additive gene effects. The importance of additive as well as non-additive gene effects with predominance of additive gene effects and inheritance of grain yield attributing traits, agreement with Saleem et al. (2010) and Saidaiah et al. (2010).
.png) Table 1 Mean square of various treatment components, estimates of components of variances for 15 different traits in 30 hybrids |
1.2 Proportional contribution
The proportional contribution of lines, testers and their interaction for the twelve quantitative and three quality traits suggested that lines played very important role for all the traits indicated dominance of parental influence in the hybrids for majority the traits. Testers were more important but their contribution was low to moderate, revealed high influence of maternal effect for all traits under study. The contribution of maternal and paternal interaction (linextester) was low to high for majority of the traits also reported by Saleem et al. (2010) and Rashid et al. (2007).
1.3 General combining ability
All the 10 lines exhibited significant and positive GCA effects for GY and tester namely PB-1 showed positive and significant GCA effects for important traits (Table 2), as valuable donors for utilizing in multiple crossing programme for obtaining higher hybrid yield through background selection. The additive gene effects were fixable in nature for grain yield and most of the yield component traits.
.png) Table 2 Estimates of general combining ability (GCA) effects of parents (10 japonica lines and 3 indica testers) for 15 characters in rice lines |
1.4 Specific combining ability effects
Out of thirty experimental hybrids five appeared as best performing, in order of merit namely TJ-16073×CSR-36, TJ-5769×NDR-359, TJ-16081×NDR-359, TJ-10365×NDR-359 and TJ-10365×PB-1 for significant and positive SCA effects due to GY and other important yield components (Table 3). The first ranking experimental hybrid TJ-16073×CSR-36 resulted highest positive and significant SCA effects for GY and contributing traits in desirable direction for PH, SF and TW. These findings were in agreement with Dalvi and Patel (2009), Salem et al. (2010) and Saidaiah et al. (2011). The second ranking experimental hybrid TJ-5769×NDR-359 showed higher positive and significant SCA effect for GY while it has significant and desirable SCA effects only for KL also noted by Allahgholipour and Ali (2006). The third ranking experimental hybrid appeared as TJ-16081×NDR-359, exhibited significant and positive SCA effects for GY and TW, KB BY also registered by Ahangar et al. (2008). Similarly, remaining two experimental hybrids TJ-16081×NDR-359 and TJ-10365×NDR-359 having significant and positive effects for GY also possessed significant SCA effects in desirable direction for other yield component, findings are agreement with Satheesh et al. (2010) and Saidaiah et al. (2011). Hence, these mentioned experimental hybrids may be considered for the development of high yielding hybrids through exploitation of hybrid vigor. The above five experimental hybrids having significant and positive SCA effects for GY and desirable significant SCA effects for SF and TW, BY and KB suggested that manifestation of SCA effects for higher GY related with higher SCA effects for important yield components. Widely the experimental hybrids exhibiting significant and desirable SCA effects were associated with better per se performance for respective traits. However, the hybrids having high SCA effects in desirable direction did not always have high mean performance for the character. Thus, the SCA effect of the experimental hybrids may not be directly related to their per se performance. This may be attributed to the fact that per se performance is a realized value whereas SCA effect is an estimate of F1 performance over parental one. Therefore, both per se performance along with SCA effects should be considered for evaluating the superiority of a desirable experimental hybrids.
.png) Table 3 Estimates of specific combining ability (SCA) effects of hybrids for 15 traits in 30 japonica × indica derived rice hybrids |
The critical evaluation revealed that experimental hybrids exhibiting high order of significant and desirable SCA effects for yield and yield contributing attributes involved parents having all types of combinations viz. high×high (H×H), high×average (H×A), low×high (L×H), average×average (A×A), average× ow (A×L), low×average (L×A) and low×low (L×L) (Table 4). The hybrids with non-significant SCA effects and parents with significant GCA effects are useful in identification of superior transgressive segregants. The observation clearly indicated no particular relationship between positive and significant SCA effects of experimental hybrids with their parents for the characters (Pratap et al., 2013).
.png) Table 4 Most promising combinations for different traits along with mean performance and gca effects of parents in 30 hybrids |
1.5 Heterosis
The mean heterosis of 30 hybrids exhibited in both positive and negative direction and varied for different characters respectively. Heterosis for GY over heterobeltiosis and standard heterosis over SV1 and SV3 were negative while mean SH over SV2 was positive (Table 5). The existence of heterosis in positive direction with expression of high degree of desirable heterosis are in conformity with Roy et al. (2009), Saidaiah et al. (2010) and Tiwari et al. (2011). The presence of high heterosis for economically important characters is not only useful for developing hybrids through exploitation of hybrid vigor, but also essential in obtaining transgressive sergeants. The estimates of heterosis for heterobeltiosis (Table 5) ranged from -75.62% (TJ-39050×NDR-359) to 40.39% (TJ-11010×PB-1) and standard heterosis varied from -80.73% (TJ-39050×PB-1) to 14.95% (TJ-11010×PB-1) over SV1, over SV2 from -65.15 (TJ-39050×PB-1) to 107.86 (TJ-11010×PB-1) and from -78.11% (TJ-39050×PB-1) to 30.61% (TJ-11010×PB-1) over SV3 respectively, agreement with Sen and Singh (2011) and Dwivedi and Pandey (2012). Out of thirty, only seven experimental hybrid namely TJ-11010×PB-1, TJ-11010×CSR-36,TJ-25892×CSR-36, TJ-25892×PB-1, TJ-16081×PB-1, TJ-10365×PB-1 and TJ-11010×NDR-359 showed significant positive heterobeltiosis (Table 6), whereas three hybrids, TJ-11010×PB-1, TJ-11010×NDR-359 and TJ-11010×CSR-36 showed positive and significant standard heterosis over SV1 (Table 7), ten hybrid combinations TJ-11010×PB-1, TJ-11010×NDR-359, TJ-11010×CSR-36, TJ-16081×NDR-359, TJ-16081×PB-1, TJ-16081×CSR-36, TJ-5769×NDR-359, TJ-10365×PB-1, TJ-10365×NDR-359 and TJ-25892×PB-1 possessed significant and positive standard heterosis over SV2 and four experimental hybrid TJ-11010×PB-1, TJ-11010×NDR-359, TJ-11010×CSR-36 and TJ-16081×NDR-359 exhibited significant and positive standard heterosis over SV3 for grain yield and some other contributing component traits (Table 7).
.png) Table 5 Mean values, performance and range of heterosis among the 30 experimental hybrids for the 15 yield related characters |
.png) Table 6 Relationship of positive heterobeltiosis for grain yield with desirable heterobeltiosis of other characters in 30 rice hybrids |
 Table 7 Relationship of positive and significant standard heterosis over SV1 (NDR-359) and SV2 (PB-1121) and SV3 (Sarjoo 52) for grain yield with standard heterosis for other characters in 30 experimental hybrids |
In this context, the most desirable hybrids showing high mean performance and highly significant heterosis of one or both types for GY were TJ-11010×PB-1, TJ-11010×NDR-359, TJ-11010×CSR-36, TJ-16081×NDR-359, TJ-16081×PB-1, TJ-5769×NDR-359, TJ-10365×PB-1, TJ-10365×NDR-359 and TJ-25892×PB-1. The hybrid TJ-11010×PB-1 showed highest mean performance for GY (34.98 g) with heterobeltiosis (40.39%) and standard heterosis over SV1 (14.95%), SV2 (107.86) and SV3 (30.61%) respectively, while highest yielding parent NDR-359, produced mean GY of 30.43 g. The other eight experimental hybrids mentioned above in order of merit, two produced higher and five lower GY than NDR-359. The best experimental hybrid TJ-11010×PB-1, produced high mean GY of 34.98g with high and significant heterobeltiosis of 40.39% with standard heterosis over SV2 (107.86%).
The other two promising hybrid namely, TJ-11010×NDR-359and TJ-11010×CSR-36 showed average SCA effect for GY as well as other characters have higher GY than NDR-359 except for the hybrid TJ-16081×PB-1 and TJ-25892×PB-1. Four experimental hybrids having positive and significant SCA effect for GY are TJ-16081×NDR-359, TJ-5769×NDR-359, TJ-10365×PB-1, and TJ-10365×NDR-359. These above six hybrids in order to merit need further testing and evaluation in adaptive trails to find out their feasibility for recommendation to use as ideal hybrid cultivars.
It was also noticed that higher heterosis over better-parent was found in some lower yielding hybrids when compared to other hybrids which have displayed high yield. This suggested that while selecting the best hybrid, besides the heterotic response over better-parent, the mean performance of the hybrids should also be given consideration. Heterosis estimate results from F1-BPH and depends more or less on the mean of the parents, There is greater possibility of getting a hybrid with lower mean performance but high heterotic response, in case the parental performance is very poor. The mean performance being the realized value and the heterotic response being an estimate, hence, weightage should be given during making selection of hybrid combinations especially when objective is to identify ideal hybrids.
1.6 Genetic distances among lines and testers
Microsatellite marker amplified a total of 131 fragments (alleles) with an average of 3.28 alleles per primer pair for 13 parental lines excluding three standard checks. The Polymorphic information content (PIC) values for the SSR loci ranged from 0.231 to 0.755 with an average of 0.542, which revealed that significant variation was present among the studied parental lines.A significant and positive correlation (r=0.350*) was found between the total number of amplified alleles and PIC values indicated, presence of broad genetic base, within lines and testers (Rabbani et al.,2010). The average GD based on the polymorphic microsatellite markers among the parental lines with testers was 0.356, ranging from 0.170 to 0.470 between GD estimates. The GD among the hybrids of PB-1 background ranged from 0.430 in the hybrid TJ–11010×PB-1 to 0.270 in TJ-39050×PB-1 with an average of 0.363 for NDR-359 GD ranged from 0.410 for hybridTJ–16081×NDR-359 to 0.310 TJ–25966×NDR-359 with an average of 0.352 for NDR-359 background. CSR-36 background resulted an average GD of 0.365 with the hybrids (0.450) TJ–25966×CSR-36 to (0.320)TJ–5769×CSR-36. Exhibited presence of significant variations among the studied parental lines. The highest GD for three different backgrounds of standard check was recorded as 0.430for hybrid TJ–11010×PB-1, 0.410 in TJ–16081×NDR-359 and (0.450) TJ–25966×CSR-36 respectively. The GDs between lines and testers spanned a great range of diversityindicating presence of broad spectrum of genetic distance and utilization in hybrid breeding programme for getting higher heterosis.
1.7 Relationships between components
Highly significant and positive association of yield and other yield contributing characters were observed with GCA but not to SCA (Table 8). The association of yield with GCA (0.964) but not to SCA (0.171) indicated biasness of selection on the basis of only GCA and unfruitfulness for exploitation of heterosis based on hybrid performance (0.637). Although GD between the parents were significantly correlated with hybrid performance, but not consistently to identify the best hybrids. This is similar to results already published (Godshalk et al., 1990; Ajmon-Marsan et al., 1998). Genetic diversity on the basis of microsatellite data set (Figure 1) and association of GD with the yield heterosis for GY surprisingly exhibited a highly significant and positive correlation of 0.663** (RH), 0.641** (BPH) and 0.638** for three SH hybrids, indicated possible utility of selected microsatellite markers for establishing strong heterotic relationship (Vuylsteke et al., 2000; Zhang et al., 2010; Singh et al., 2011).
 Table 8 Association of yield attributing traits with genetic components and heterosis in 30 hybrids |
 Figure 1 Amplification pattern of the marker HRM 15630 among the rice lines analyzed with Nipponbar (japonica) and jyothi (indica)as reference |
1.8 Wide combining ability analysis
A major difficulty encountered in the development of inter-specific hybrids is the partial hybrid sterility and panicle emergence, frequently has been observed in most japonica×indica hybrids (Kato et al., 1928). A special class of rice germplasms, have been identified by many workers and referred as wide-compatible varieties, capable to contribute towards enhanced spikelet fertility in hybrids than the normal fertility when crossed with japonica and indica groups (Ikehasi and Araki, 1984). Out of 16 lines including Nagina-22 as wide compatible indica, jyothi as normal indica and Nipponbare as japonica reference, 11 lines amplified 417-bp fragment in indica and japonica alleles including Nipponbare, Jyothi along with three testers viz. PB-1, NDR359, CSR-36 and 6 japonica groups namely, TJ-5769,TJ-64897, TJ-16081, TJ-0365, TJ-4122 and TJ-25892 (Figure 2). A 281-bp fragment were identified to possess neutral allele in Nipponbare, Jyothi, Nagina22 (Reference) and 7 japonica lines as TJ-5769, TJ-64897, TJ-25966, TJ-1010, TJ-16081, TJ-17073 and TJ-39050 at S5 locus. These lines could be utilized in inter specific hybridization to exploit higher level of heterosis. Although, Jyothi and four japonica group namely, Nipponbare, TJ–5769, TJ–64897 and TJ–16081 exhibitedheterozygous nature possessing both type of bands. The present study clearly demonstrated the utility of primer S5 InDel in identifying wide compatible genotypes possessing S5 neutral allele(Sundaram et al., 2010).
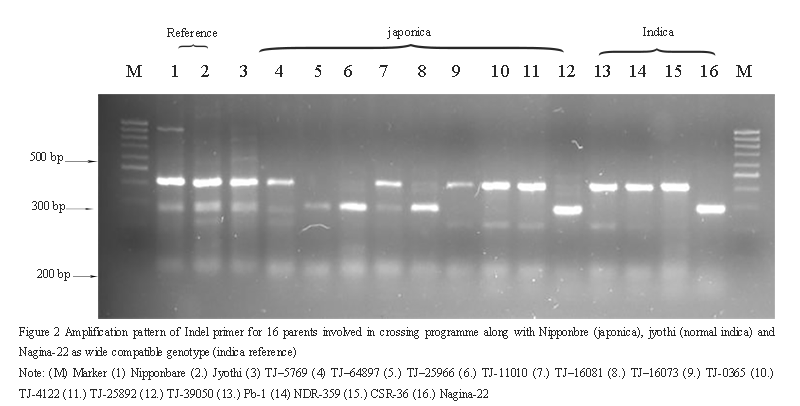 Figure 2 Amplification pattern of Indel primer for 16 parents involved in crossing programme along with Nipponbre (japonica), jyothi (normal indica) and Nagina-22 as wide compatible genotype (indica reference) |
2 Discussion
Designed experiment revealed that both per se performance of parents involve in hybridization programme along with the derived hybrids having high SCA effects should be considered for evaluation of superior desirable experimental hybrids. Higher GCA variances than corresponding SCA indicated preponderance of additive gene effects for maintaining heterozygosity for improving desirable traits. The selection of the most promising experimental hybrids should be based on ony GCA but also per-se performance and SCA.The hybrids having higher heterosis over heterobeltiosis in some lower yielding hybrids, as compared to high yielding hybrids suggested, best hybrid selection, besides the heterotic response over better-parent, mean performance of the hybrids. Heterosis estimate results of heterobeltiosis depends more or less on the mean of the parents in question. There is possibility of getting a good high yielding hybrid with lower mean performance but heterotic response of the parent may be very poor due to their genetic makeup and diversity. Highly significant and positive correlation of yield with GCA but not to SCA indicated biased selection and will not be worthwhile for exploitation of heterosis. Wide combining ability study revealed that the genotypes having natural allele at S5 locus can give rise to fully fertile panicle as good contributer towards yield. The presence of desirable neutral allele possessing genotype can be involve for getting the higher level of yield heterosis in japonica and indicia hybrids combinations.
3 Materials and Methods
3.1 Plant Martials
The parental material for the present study comprised of 10 japonica lines namely, TJ–5769, TJ–64897,TJ–25966,TJ-11010, TJ–16081, TJ–16073, TJ-0365, TJ-4122, TJ-25892, TJ-39050 and three indica testers NDR-359, CSR-36 and PB-1 were used to derive 30 F1’s (Figure 1) following the Line×Tester fashion design during Kharif (2010) and resultant of 30 F1s along with 13 parents were evaluated in randomized complete block design (RCBD) with three replications and NDR-359, PB-1121 and Sarjoo-52 national check during kharif (2011) at Crop Research Farm, Nawabganj, C.S. Azad University of Agriculture and Technology, Kanpur at the gangetic plain of central Uttar Pradesh, India. The site of experiment was salt affected clay loam (natrustalf) soil having pH=10.5, EC=2.03 and low in organic carbon, nitrogen and phosphorus. Observations were recorded on ten competitive plants randomly selected in each replications for twelve quantitative and three quality traits viz., days to 50% flowering (DF), plant height (PH), panicle bearing tillers per plant (PBT), flag leaf area (FLA), panicle length (PL), panicle weight (PW), spikelets per panicle (SP), spikelet fertility (SF), 1000-grains weight (TW), biological yield per plant (BYP), harvest index (HI), grain yield per plant(GY), kernel length (KL), kernel breadth (KB) and L:B ratio (LBR).
3.2 Wide compatibility approaches
Generally large number of hybrid combinations is required for selecting few good hybrids. It requires alternative approaches to enhance the selection efficiency and to asses hybrid performance. Refining these difficulties a set of INDEL primers were designed and utilized for screening the neutral allele possessing the high spikelet fertility and contribution in yield enhancement. A total of 16 parents (including Nagina-22 as wide compatible indica reference, jyothi as normal indica and Nipponbare as japonica reference) were raised in pots and total genomic DNA was isolated from 25 days young leaves following CTAB extraction method (Murray and Thompson, 1980) with some modification. Availability of molecular markers, fine genetic maps and genome sequences in rice provided the opportunity for successful cloning of major S5 QTLs (Chen et al., 2008). A discontinuous stretch of 136 bp deletion spread by TATA motifs in first exon of the gene coding for aspartic protease was noticed in wide compatible varieties carrying the neutral allele, compared to the japonica and indica alleles (Ikehashi and Araki, 1986). The large insertion/deletion (Indel) provided opportunity for utilizing as PCR based marker for screening wide-compatible genotypes. Sundaram et al. (2010) developed a PCR-based multiplex marker system (S5-MMS) targeting 136 bp deletion of S5 neutral allele, suitable for identification of allelic status at the major wide compatibility gene locus S5 in rice germplasm. The wide compatibility e study was subjected to molecular amplification with InDel primers developed by Sundaram et al. (2010).
3.3 Statistical analysis
The adjusted mean data of different traits was subjected to analysis by standard statistical and biometrical method for Line×Tester analysis by Kempthorne (1957), General combining ability (GCA) and Specific combining ability (SCA) effects for grain yield and other yield contributing characters, were calculated according to the line×tester model. The Relative heterosis (RH), heterobeltiosis (BPH) and standard heterosis (SH) of each cross was calculated as the difference between F1 means and the respective mid-parent, better-parent and over three standard variety SV1 (NDR-359) and SV2 (PB-1121) and SV3 (Sarjoo-52)by Fonseca and Patterson (1968). Association approach for relationship between grain yield (GY) and general combining ability (GCA), specific combining ability (SCA), molecular markers amplification based genetic diversity (GD) with hybrid yield, relative heterosis (RH), better parent heterosis (BPH) and standard heterosis (SH) was calculated according to Salvaraj et al. (2010).
3.4 Molecular marker assay and data analysis
10 japonica lines and three indica testers namely, NDR-359, CSR-36 and PB-1 along with Nagina-22 as wide compatible indica, Jyothi as normal indica and Nipponbare as japonica reference were subjected to PCR amplification for analysis of molecular genetic diversity with 40 uniformly distributed hypervariable microsatellite markers (Soni et al., 2013). InDel PCR based markers targeting 136 bp deletion at S5 locus to discriminate the presence of neutral allele (281 bp) were utilized for screening of neutral allele designed by Sundaram et al. (2010) along with following component mixture of a 10 µl volume for each PCR reaction: each primer 0.25 µM, 10x PCR buffer with 1.5 mM MgCl2, 2.5 mM dNTPs, 1U Taq DNA polymerase (Invetrogen, USA) and approximately 25-30 ng of tempelet DNA. The touchdown PCR profile was: initial denaturation at 95°C for 5 min followed by 12 cycles of deneturation for 30 s at 94°C, 30 s annealing at 72°C~63°C (1°C decrease per cycle), 1 min extension at 72°C and 25 cycles of denaturation for 30 s at 94°C, 30 s annealing at 62°C, I min extension at 72°C, with a final extension of 72°C for 7 min. The amplified PCR products were separated on a 1.5% Seakem® LE agarose gel (Loanza, USA), stained with ethidium bromide and visualised under a UV transilluminator (Alpha Innotech, USA). Gel was scored base on the banding pattern as, indica/japonica type (417-bp), neutral (281-bp) and heterozygous (both) alleleand genetic diversity was calculated by reducing similarity value from unity Vuylsteke et al. (2000) and Soni et al. (2013).
Acknowledgments
Author would like to thank Dr. R.M. Sundaram (Principal Scientist) Biotechnology, Dr. V.P. Bhadana (Principal Scientist) Department of Genetics and Plant Breeding, Hybrid Rice Section, Indian Institute of Rice Research, Rajendranagar, Hyderabad (Telangana) India, for providing the materials and facility for carry out molecular experiments and Dr. V.K. Yadav (Rice Breeder) Department of Genetics and Plant Breeding, CSAUA&T, Kanpur (UP) India for conducting the field trials timely, valuable suggestions and completion of this manuscript.
Ahangar L., Ranjbar G.A., and Nouroozi M., 2008, Estimates of combining ability for yield and yield components in rice (Oryza sativa L.) cultivars using diallel cross, Pakistan Journal of Biological Sciences, 11(9): 1278-1281
http://dx.doi.org/10.3923/pjbs.2008.1278.1281
Ajmone M.P., Castiglioni P., Fusari F., Kuiper M., and Motto M., 1998, Genetic diversity and its relationship to hybrid performance in maize as revealed by RFLP and AFLP markers, Theoretical & Applied Genetics, 96(2): 219-227
http://dx.doi.org/10.1007/s001220050730
Allahghlilpour M., and Ali A.J., 2006, Gene action and combining ability for grain yield and its components in rice, Journal of Sustainable Agriculture, 28(3): 39-53
http://dx.doi.org/10.1300/J064v28n03_05
Chen J., Ding J., Ouyang Y., Du H., Yang J., Cheng K., Zhao J., Qiu S., Zhang X., Yao J., Liu K., Wang L., Xu C., Li X., Xue Y., Xia M., Ji Q., Lu J., Xu M., and Zhang Q., 2008, A triallelic system of S5 is a major regulator of the reproductive barrier and compatibility of indica-japonica hybrids in rice, Proceedings of the National Academy of Sciences USA, 105(32): 11436-11441
http://dx.doi.org/10.1073/pnas.0804761105
Dalvi V.V., and Patel D.U., 2009, Combining ability analysis for yield in hybrid rice, Oryz- An International Journal on Rice, 46(2): 97-102
Dwevedi D.K., and Pandey M.P., 2012, Gene action and heterosis for yield and associated traits in Indica and Tropical japonica crosses of rice (Oryza sativa L.) involving wide compatibility gene(s), International Journal of Plant Breeding and Genetics, 6(3): 14-150
http://dx.doi.org/10.3923/ijpbg.2012.140.150
Fonseca S., and Patterson F.L., 1968, Hybrid vigor in seven parent diallel crosses in Common Wheat (T. aestivum L.), Crop Science, 8(1): 85-88
http://dx.doi.org/10.2135/cropsci1968.0011183X000800010025x
Godshalk E.B., Lee M., and Lamkey K.R., 1990, Relationship of restriction fragment length polymorphism to single cross hybrid performance of maize, Theor. Appl. Genet., 80(2): 273-280
http://dx.doi.org/10.1007/BF00224398
Ikehashi H., and Araki H., 1984, Variety screening of compatibility types revealed in F1 fertility of distant cross in rice, Japanese Journal of Breeding, 34(3): 304-313
http://dx.doi.org/10.1270/jsbbs1951.34.304
Ikehashi H., and Araki H., 1986, Genetics of F1 sterility in remote crosses of rice, Rice Genetics, pp.119-130
Kato S., Kosaka H., and Hara S., 1928, On the affinity of rice varieties as shown by fertility of hybrid plants, Bull Sci Fac Agric Kyushu Univ, 3: 132-147
Kempthrorne O., 1957, An introduction to Genetical Statistics, John Wiley and Sons Inc, New York., pp.468-471
Khush G.S., and Virk P.S., 2000, Rice breeding achievements and future strategies, Crop Improvement, 27(2): 115-144
Khush G. S., 2013, Strategies for increasing the yield potential of cereals: case of rice as an example, Plant Breeding, 132(5): 433-436
http://dx.doi.org/10.1111/pbr.1991
Khush G.S., and Brar D.S., 2002, Biotechnology for rice breeding: progress and impact, In: Sustainable rice production for food security, Proceedings of the 20th Session of the International Rice Commission, Bangkok, Thailand, pp.23-26
Murray M.G., and Thompson W.F., 1980, Rapid isolation of high molecular weight plant DNA, Nucleic Acids Research, 8(19): 4321-4325
http://dx.doi.org/10.1093/nar/8.19.4321
Pratap N., Shekhar R., Singh P.K., and Soni S.K., 2013, Combining Ability, Gene Action and Heterosis Using CMS Lines In Hybrid Rice (Oryza sativa L.), The Bioscan, 8(4): 1521-1528
Rabbani M.A., Masood M.S., Shinwari Z.K., and Shinozaki K.Y., 2010, Genetic analysis of basmati and non-basmati Pakistani rice (Oryza sativa L.) cultivars using microsatellite markers, Pakistan Journal of Botany, 42(4): 2551-2564
Rashid M., Cheema A.A., and Ashraf M., 2007, Line × tester analysis in Basmati rice, Pakistan Journal of Botany, 36(6): 2035-2042
Roy S.S., Senapati B.K., Sinhamahapatra S.P., and Sarkar K.K., 2009, Heterosis for yield and quality traits in rice, Oryz- An International Journal on Rice, 46(2): 87-93
Saidaiah P., Ramesha M.S., Kumar S.S. and Suresh J., 2011, Combining ability analysis for yield and yield component traits in rice (Oryza sativa L.), Progressive Agriculture, 11(2): 293-297
Saidaiah P., Sudheer K.S., and Ramesha M.S., 2010, Combining ability studies for development of new hybrids in rice over environments, Journal of Agricultural Science, 2(2): 225-233
http://dx.doi.org/10.5539/jas.v2n2p225
Saleem M.Y., Mirza J.I., and Haq M.A., 2010, Combining ability analysis for yield and related traits in Basmati rice (Oryza sativa L.), Pakistan Journal of Botany, 42(42): 627-637
Satheesh K.P. and Saravanan S.T., 2010, Combining ability for yield and yield contributing characters in rice (Oryza sativa L.), Electronic Journal of Plant Breeding, 1(5): 1290-1293
Selvaraj I., Nagarajan P., Thiyagarajan K., and Bharathi M., 2015, Predicting the relationship between molecular marker heterozygosity and hybrid performance using RAPD markers, African Journal of Biotechnology, 9(45): 7641-7653
Sen C., and Singh R.P., 2011, Study on heterosis in Boro × High yielding rice hybrids, International Journal of Plant Breeding and Genetics, 5(2): 141-149
http://dx.doi.org/10.3923/ijpbg.2011.141.149
Singh V.K., Upadhyay P., Sinha P., Mall A.K.., Kumar R., Singh E.A., Jaiswal S.K., Biradar S., Ramakrishna S., Sundaram R.M., Ahmed I., Viraktamath B.C., Kole C., and Singh S., 2011, Prediction of hybrid performance based on the genetic distance of parental lines in two-line rice (Oryza sativa L.) hybrids, Journal of Crop Science & Biotechnology, 14(1):1-10
http://dx.doi.org/10.1007/s12892-010-0111-y
Soni S., Sundaram R.M., Yadav V.K, and Bhadana V.P., 2013a, Genotyping of world mini core collections, their unique identification and genetic similarity using microsatellite markers in rice (oryza sativa l.), Phillippines, pp.377
Soni S, Sundaram R.M., Yadav V.K., and Bhadana V.P., 2013b, Development of Key marker concept for prediction and validation of heterosis using microsatellite markers in parental lines of rice (Oryza sativa), 7th International Rice Genetics Symposium,Phillippines, pp.376
Sundaram R.M., Saktivel K., Hariprasad A.S., Ramesha M.S., Viraktamath B.C., Neeraja C.N., Balachandran S.M., Sobha Rani N., Revathi P., Sandhya P., and Hari Y., 2010, Development and validation of a PCR based functional marker system for the major wide compatible genes locus S5 in rice, Molecular Breeding, 26(4): 719-727
http://dx.doi.org/10.1007/s11032-010-9482-5
Tiwari D.K., Pandey P., Giri S.P., and Dwivedi J.L., 2011, Prediction of genetic action, heterosis and combining ability to identify superior rice hybrids, International Journal of Botany, 7(2): 126-144
http://dx.doi.org/10.3923/ijb.2011.126.144
Vuylsteke M., Kuiper M., and Stam P., 2000, Chromosomal regions involved in hybrid performance and heterosis: their AFLP-based identification and practical use in prediction models, Heredity, 3(3): 208-218
http://dx.doi.org/10.1046/j.1365-2540.2000.00747.x
Zhang T., Ni X.L., Jiang K.F., Yang Q., Yang L., Wan X.Q., Ying J., and Zheng J.K., 2010, Correlation between heterosis and genetic distance based on molecular markers of functional genes in rice, Rice Science, 17(4): 13-20
http://dx.doi.org/10.1016/S1672-6308(09)60029-9



. PDF(310KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Soni Kumar
. Yadav Kumar
. Bhadana Pal
. Yadav Chand
. Sundaram Minakshi
Related articles
. Wide compatibility
. Microsatellite markers
. Economic heterosis
. Association study
. Neutral allele
. Higher yield
Tools
. Email to a friend
. Post a comment