Research Report
Marker-assisted Introgression of a Broad-spectrum Resistance Gene, Pi40 Improved Blast Resistance of Two Elite Rice (Oryza sativa L.) Cultivars of Turkey 
2 Novel Gene Resources Laboratory, Plant Breeding Division, International Rice Research Institute, DAPO Box 7777, 1226 Metro Manila, Philippines
3 Trakya Agricultural Research Institute, P. Box 16, 22100, Edirne, Turkey
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2016, Vol. 7, No. 33 doi: 10.5376/mpb.2016.07.0033
Received: 16 May, 2016 Accepted: 01 Jul., 2016 Published: 18 Sep., 2016
Beşer N., Del Valle M.M., Kim S.M., Vinarao R.B., Sürek H., and Jena K.K., 2016, Marker-assisted Introgression of a Broad-spectrum Resistance Gene, Pi40 Improved Blast Resistance of Two Elite Rice (Oryza sativa L.) Cultivars of Turkey, Molecular Plant Breeding, 7(33): 1-15 (doi: 10.5376/mpb.2016.07.0033)
Rice blast, caused by Magnaporthe oryzae is the most serious fungal disease of cultivated rice (Oryza sativa). In serious disease infection, significant yield loss in japonica and indica rice occurs every year worldwide. Of the 100 blast resistance genes identified, majorities are race specific and resistance breaks down rapidly. However, the new resistance gene, Pi40 shows broad-spectrum resistance to blast races in many tropical and temperate rice growing countries including Turkey. In this study, the Pi40 gene was introgressed into two Turkish elite cultivars, Osmancik-97 and Halilbey, to improve with broad-spectrum durable blast resistance and high yield potential through marker-assisted backcross breeding. Advanced backcross lines (ABLs) of BC3F6 and BC4F6 generations were produced from the crosses of Osmancik-97 and Halilbey with the Pi40-donor line, IR83260-1-1-1-5-B-3-1-2-B. ABLs with homozygous Pi40 alleles were selected by foreground analysis using gene-specific CAPS DNA marker 9871.T7E2b. The two varieties were expressing high susceptibility to blast disease isolates in Turkey and Philippines. Eleven selected ABLs expressed strong resistance to blast races in natural field and spray inoculation conditions. Background analysis of selected ABLs with 6K SNP assays showed 90.07% and 70.78% recovery of the recurrent genotype for Osmancik-97 and Halilbey ABLs respectively. We have produced valuable resources for blast disease resistance and developed improved breeding lines with strong resistance to blast for their cultivation in Turkey. Our new strategy of transferring the novel blast resistance gene (Pi40) into Turkish varieties will enhance rice production and productivity in the temperate regions of the world.
Introduction
Rice (Oryza sativa L.) is the staple food for more than half of the world population. Increasing global population escalates demand for rice consumption particularly in most of the Asian countries. In this context, there is a need to increase rice production more than 40 percent by 2030 to feed the increasing population (Khush, 2005). Even though rice ranks second to wheat in terms of area harvested; but in terms of importance as food crop, rice provides more calories per hectare than any other cereal crop (De Datta, 1981). Rice is cultivated in tropical as well as temperate ecosystems as indica and japonica cultivars respectively. Rice cultivated in limited areas of temperate regions of Turkey is a typical japonica type where rice consumption is increasing (Beser and Surek, 2012).
In a modernised agriculture system, major biotic and abiotic stresses limit rice production due to the emergence of new pathotypes of diseases, breakdown of resistance genes in elite cultivars and climatic changes (Normile, 2008). Of the several biotic stresses, blast disease causes significant yield loss annually. Rice blast caused by the fungus Magnaporthe oryzae (anamorph: Pyricularia grisea Sacc.) is generally considered as the most severe disease worldwide due to its widespread distribution in over 80 countries and becomes epidemic in severe infections (Ou, 1980; Zeigler et al., 1994).
Turkey is one of the most favoured countries in terms of agricultural production due to good ecological and climatological conditions, land property and a rich crop pattern. The main rice growing regions in Turkey are the North western (Marmara-Thrace) and the northern parts of the country (Black sea region). Blast disease is one of the most serious problems in temperate rice cultivation of Turkey. A heavy blast epidemic occurred in some growing areas at northwest of Turkey in 1995 and 1997 which caused 25 percent yield loss in 25,000 hectares of rice growing areas in the region. Therefore, varietal improvement for blast disease resistance is imperative toward development of high yielding varieties (Beşer and Sürek, 2012).
Osmancık-97 is a semi-dwarf elite Turkish cultivar released in 1997. It has all desirable agronomic and grain quality traits with a high milling yield potential but susceptible to blast disease (Manners, 2013). Halilbey is another elite variety with high yield potential but highly susceptible to leaf and panicle blast. In some years, rice production and rice quality is reduced due to severity of the blast disease. Therefore, the development of a blast-resistant cultivar has still been the most effective and economical breeding strategy to control the spread of the disease (Jeung et al., 2007). Resistance of rice cultivars to blast disease are developed by conventional breeding using the sources of resistance from land races or diverse rice varieties. However, the resistance breaks down in two to three years (Miah et al., 2013). Novel sources of blast resistance genes have been identified in the introgression lines derived from wild Oryza species (Jena and Kim, 2010). Of the 100 blast resistance genes, the Pi40 gene has been identified in an indica introgression line that inherited the resistance gene from a wild species O. australiensis and has durable broad-spectrum of blast resistance (Jeung et al., 2007; Sharma et al., 2012; Suh et al., 2009). The Pi40 gene confers blast resistance at seedling as well as reproductive stages. It has shown resistance to emerging virulent blast strains in several countries and several provinces of China (Wang, 2012,). Marker-assisted backcross breeding is a powerful tool to incorporate novel resistance genes into susceptible cultivars and develop improved breeding lines in a short period of time (Jena and Mackill, 2008).
The objective of this study is to incorporate the Pi40 gene into two Turkish elite cultivars, Osmancık-97 and Halilbey, through marker-assisted backcross breeding and develop improved blast resistant cultivars for Turkey. This study will lead to the production of promising breeding lines with blast resistance in a short period of time using resistance gene specific molecular marker analysis tools. Background genome analysis of progenies will show conversion of recurrent parent genome with the Pi40 gene-derived blast resistance introgression.
1 Results and Analysis
1.1 Development and molecular analysis of ABLs of Osmancık-97and Halilbey
Five F1 plants from the crosses of Osmancık-97 and Halilbey with donor parent (DP) were produced. Hybridity of the F1 plants were confirmed and were further backcrossed to recurrent parent (RP). The presence of the Pi40 gene was tracked in backcross (BC) progenies by marker-assisted selection (MAS). Several BC progenies from Osmancık-97 and Halilbey crosses were selected and validated by MAS (Figure 1a). Selected plants having the Pi40 gene were advanced until BC3F5.6 and BC4F5.6 and the progenies having phenotypic traits similar to the respective RP were selected (Supplementary Figure 1).
|
Figure 1a Foreground analysis of advance backcross lines (ABLs) of Osmancik-97. DNA was amplified with 9871.T7E2b as a primer for the Pi40 gene. Arrows indicate resistance specific-alleles. Legend: Kb+= DNA ladder; P1=Osmanc?k-97; P2=IR83260-1-1-1-5-B-3-1-2-B; 1-1=ABLs |

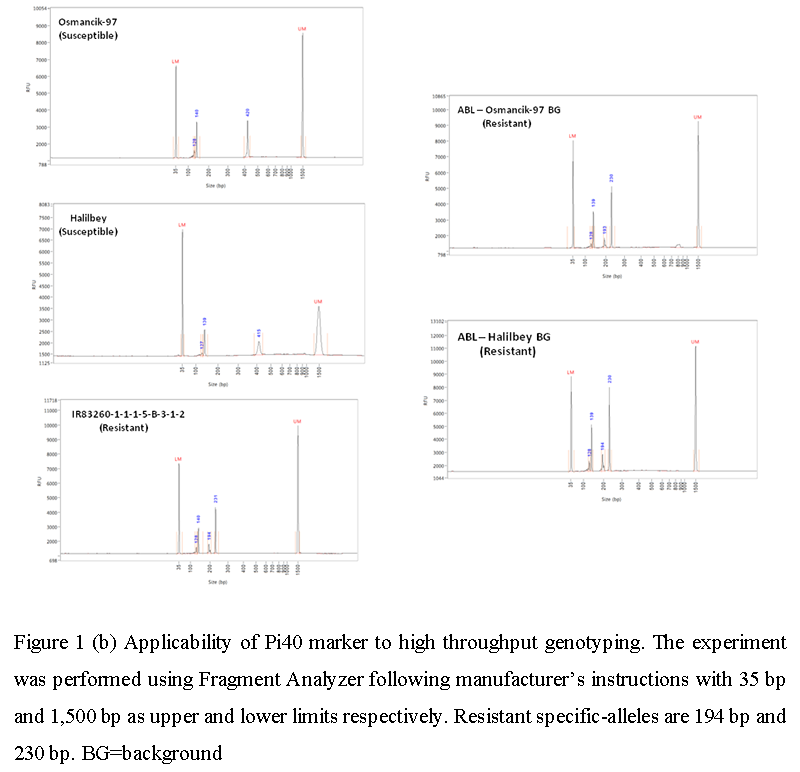
The usability of Pi40 gene-specific marker in high throughput genotyping technologies was checked using Fragment Analyzer. Capillary electrophoresis of polymerase chain reaction (PCR) amplicons of the susceptible recurrent parents detected three susceptible allele-specific bands, 128 bp, 140 bp, and 415 bp for Halilbey, and 128 bp, 140 bp, and 420 bp for Osmancik-97. On the other hand, four bands were detected in the resistant genotype, donor parent. Among the four products, two amplicons (resistance specific-alleles) were unique to the donor parents which were 194 bp and 230 bp. Eleven advanced backcross lines (ABLs) were also analyzed and all showed the presence of the resistance specific-alleles (Figure 1b). The designed DNA marker linked to the Pi40 gene required the digestion of the PCR products with MluC1 which produced two unique bands for the resistant allele while the susceptible allele showed two common bands. Our results from the Fragment Analyzer were in congruence with these as it was able to detect the same profile of bands in susceptible and resistant genotypes (Figure 1b).
|
Figure 1b Applicability of Pi40 marker to high throughput genotyping. The experiment was performed using Fragment Analyzer following manufacturer’s instructions with 35 bp and 1,500 bp as upper and lower limits respectively. Resistant specific-alleles are 194 bp and 230 bp. BG=background |
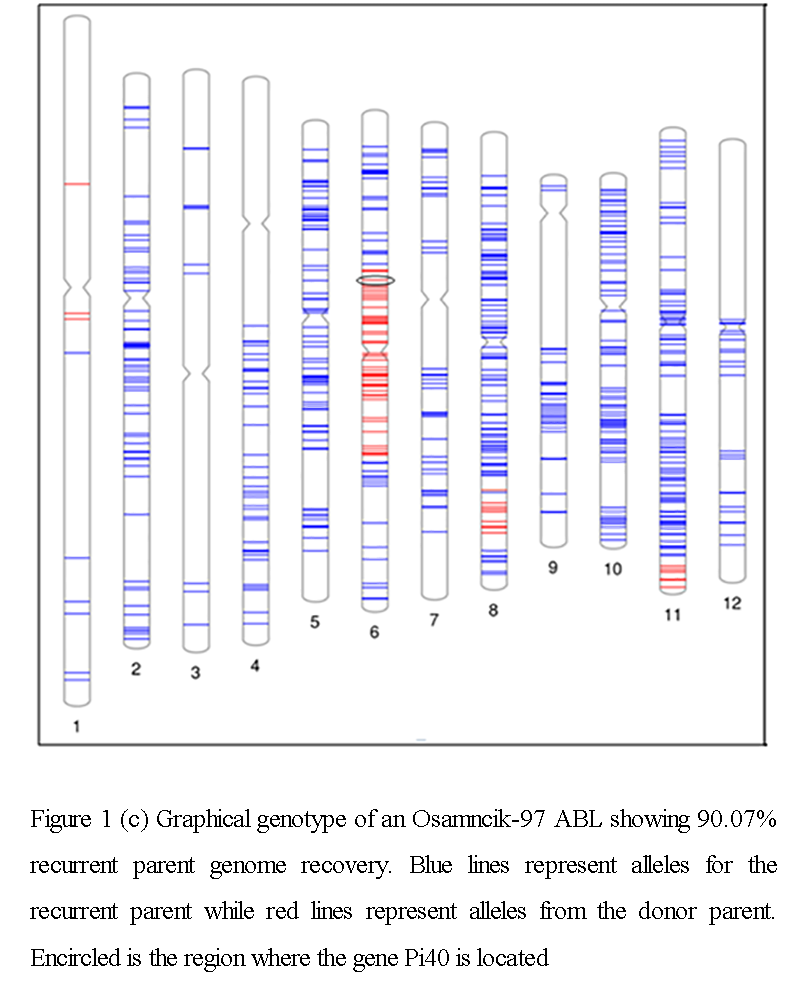
A total of 14 lines were processed for background genotyping which includes 11 ABLs, two recurrent parents (Halilbey and Osmancik-97), and the DP by Illumina 6K Infinium single nucleotide polymorphism (SNP) Chip from Cornell University was used in this study and was originally designed to contain a total of 5,274 SNPs. After employing quality controls and clustering, a total of 4,606 SNPs were found to be of high quality. Comparing the SNP genotypes of Halilbey and DP, 795 (17.26%) of the SNPs were found to be polymorphic between the parents. These markers were distributed across the 12 chromosomes of rice with an average distance of 0.44 Mb between markers. In case of Osmancik-97 and DP, 676 (14.68%) SNPs were found to be polymorphic providing an average genomic coverage of 0.43 Mb interval. A total of four ABLs of Osmancik-97 were SNP genotyped. These lines had an average recurrent parent genome recovery of 88.75% and ranges from 87.7 to 90.07% (Figure 1c). A total of seven ABLs were genotyped from the cross between Halilbey and DP. These ABLs registered an average background genome recovery of 66.88%, which ranges from 63.98 to 70.78% (Supplementary Figure 2). SNP markers designed in certain regions of chromosomes 1, 3, 4, and 12 were not polymorphic between Osmancik-97 and DP which left these regions partly characterized or uncharacterized at all. Further analysis of these regions will be carried out using simple sequence repeat markers.
|
Figure 1c Graphical genotype of an Osamncik-97 ABL showing 90.07% recurrent parent genome recovery. Blue lines represent alleles for the recurrent parent while red lines represent alleles from the donor parent. Encircled is the region where the gene Pi40 is located. |
1.2 Evaluation for blast resistance
Selected ABLs of Halilbey and Osmancik-97 were evaluated for blast resistance in uniform blast nursery (UBN) during 2014WS and 2015DS. Halilbey and Osmancik-97 showed susceptibility to blast during the 2014WS evaluation. Both cultivars recorded scores of 6 and 5 for Halilbey and Osmancik-97 respectively. The DP and resistant (R) check were resistant with a score of 1. Scores of the seven ABLs of Halilbey consistently showed resistance with a mean of 0.43 score. Likewise, Osmancik-97-derived ABLs also showed resistance to blast under natural field conditions registering mean score of 1.0. Percent disease leaf area (DLA) of the selected lines were also recorded and were observed to be of congruence to the results using standard evaluation system (SES) of scoring. Halilbey-derived ABLs recorded an average DLA of 0.21% and ranges from 0% to 0.5%. The same observation was also recorded in ABLs of Osmancik-97 which showed resistance with DLA of 0.38%. The same experiment was carried out during 2015DS. Results obtained from this experiment produced results that were similar to the 2014WS evaluation (Figure 2a, Figure 2b).
|
Figure 2 Screening for blast resistance under field and glasshouse conditions. (a) Disease scores of advance backcross lines (ABLs) and parents using Standard Evaluation System of IRRI and (b) Percent diseased leaf area of ABLs and parents. (c) Blast disease reaction in glasshouse of ABLs and parents using strain M101-1-2-9-1. Legend: 1=Pi40; 2=LTH, 3=CO39, 4=Osmancik-97, 5=Halilbey; 6=IR83260-1-1-1-5-B-3-1-2-B, 7-14=ABLs. |
.png)
Twenty one different virulent strains of Philippines were used to evaluate parents and ABLs for blast reaction by spray inoculation method in glasshouse. Seven ABLs derived from Halilbey were tested. Of these ABLs, one (ABL9) showed resistance to all of the strains tested. On the average, Halilbey-derived ABLs showed resistance to 18 out of 21 (85.7%) isolates tested while Halilbey showed resistance to only four (19%) strains (Figure 2c). All ABLs derived from Halilbey showed resistance to strains M36-1-3-10-10, JMB840610, Ca23-49, V86010, M39-1-2-21-2, M39-1-2-21-2, 9482-01-03, JMB8401, Ca41, and Pi9-G7-2K-1 while Halilbey showed susceptibility to these strains. Osmancik-97 showed resistance to seven strains (33.3%) while its progenies showed average resistance to 19 strains (90.5%) and ranged from 17 to 21 (81% to 100%). Progenies from Osmancik-97 showed resistance to M36-1-3-10-10, Ca23-49, M39-1-2-21-2, BN111, 9482-01-03, IK81-3, and B90002 for which Osmancik-97 was susceptible (Table 1). ABL1 to ABL11 were also evaluated for field blast resistance in Turkey. The RPs, Osmancik-97 and Halilbey, were susceptible while all the selected ABLs were resistant (Table 2). These results clearly showed the production of improved breeding lines from susceptible RPs by introgressing the Pi40 gene for blast resistance.
|
Table 1 Blast disease reaction of selected advance backcross lines to blast strains using spray inoculation method and field blast screening test Note:*Blast disease scoring following JIRCAS, Japan scale: R=Resistant (0) no visible lesion; (1) needle-like lesion; (2) round circular lesion without gray center; (3) round circular lesion with gray centre; (4) spindle type lesion with gray center (5) merged spindle type lesion with gray center; Strains (1-23) used were 1=M36-1-3-10-10; 2=JMB840610; 3=Ca23-49; 4=V850256; 5=V86010; 6=M39-1-3-B-1; 7=M39-1-2-21-2; 8=Ca89; 9=BN209; 10=BN111; 11=43; 12=M101-1-2-9-1; 13=9482-01-03; 14=IK81-3; 15=JMB8401; 16=Ca41; 17=IK81-25; 18=P06-06; 19=B90002; 20=M64-1-3-9-1; 21=Pi9-G7-2K-1; ABL=Advanced Backcross Lines; BG=background; Os=Osmancik-97; Hal=Halilbey; RP=Recurrent Parent; 4128=IR83260-1-1-1-5-B-3-1-2-B; Gen=Generation; ND=No Data; FBST=Field Blast Screening Test at Trakya, Turkey; R=Resistant; S=Susceptible |
.png)
|
Table 2 Agronomic traits of selected advance backcross lines of Osmancik-97 and Halilbey Note: *=mean value significantly different with respective recurrent parent; aSignificance calculated at p<0.05; RP= recurrent parent; DTH=days to heading; PH=plant height; TN=tiller number; FLL=flag leaf length; 100GW=hundred grain weight; PL=panicle length; GN= grain number; F=spikelet fertility; BG=background; Gen=generation; Hal=Halilbey; Os=Osmancik-97 |
.png)
2 Discussion
Host-plant resistance is the most important breeding strategy to breed cultivars with durable resistance to blast disease of rice (Khush and Jena, 2009). Some cultivars with major resistance genes (Pi2 and Pi9) are cultivated for a long period of time without loss of resistance to blast (Bonman et al., 1992; Bonman and Mackill, 1988; Jeung et al., 2007). Additionally, a novel resistance gene, Pi40 has been tested for its durability of blast resistance and the gene has shown promise for durable resistance that is being used in several rice breeding programs (Suh et al., 2009).
Although a number of resistance genes have been identified and used in the breeding program, most of those genes breakdown within a short period (Fukuta et al., 2009). Wild species of rice are reservoir of many valuable genes including blast resistance which could be exploited for improving elite cultivars (Jeung et al., 2007). One of the several identified genes, Pi40 is derived from the wild species, O. australiensis (Acc. 100882) of EE genome and reported to have resistance to a diverse group of M. grisea isolates (Suh et al., 2009).
In this study, the transfer of the durable blast resistance gene, Pi40 into two Turkish elite cultivars using marker-assisted backcross breeding has been considered as an effective approach. Several blast resistance genes including Pi-b, Pi-k, Pi-i, Pi-z, and Pi-ta, have also been introgressed in different breeding programs using similar approach (Miah et al., 2013). In case of Pi40, an earlier work also introgressed the gene in two japonica cultivars, Jinbubyeo and Junambyeo (Suh et al., 2009). Inter-varietal cross progenies were produced using Osmancık-97 and Halilbey as recurrent parents and transferred the Pi40 gene from a japonica genetic background. BC3F5.6 and BC4F5.6 progenies were developed by repeated backcrossing to the respective recurrent parents after conducting foreground selection for the target Pi40 gene. Further confirmation of the selected plants at each backcross generation has been carried out by blast disease evaluation in natural field and spray inoculation conditions. This approach has enhanced the selection efficiency for the target blast resistance gene at a shortest time possible as it has been shown in case of transferring brown planthopper (BPH) resistance gene, Bph18 and submergence tolerance gene, Sub1 in rice (Neeraja et al., 2007; Suh et al., 2011).
The Pi40 gene conferred blast resistance to virulent strains in the field as well as selected virulent blast strains in the glasshouse. ABLs tested showed resistance to majority of the blast isolates used. However, some ABLs (ABL2, ABL3, and ABL4) showed susceptibility to strain P06-06 even though the donor and recurrent parents were resistant. This result suggested the loss of resistance gene during meiotic recombination and it was similar to the findings of Fu et al., (2012). The identification of the Pi40 gene in the selected blast resistant plants from different backcross generations by foreground analysis revealed the power of the DNA marker, 9871.T7E2b which has been designed from NBS-LRR motif sequences (Jeung et al., 2007). This study not only revealed the successful transfer of blast resistance gene into Osmancık-97 and Halilbey but also confirmed tight linkage between the Pi40 gene specific marker and blast resistance phenotype. Interestingly, the early generation lines selected for blast resistance expresses resistance to blast races/strains in Turkey in a preliminary field test. Also, further testing of advanced backcross generation progenies at several locations in Turkey revealed that all ABLs we selected confirmed resistance to blast. This study will enable the isolation of promising breeding lines for their adaptation and cultivar development.
The major target in this study was the introgression of the Pi40 gene from the donor parent (IR83260-1-1-1-5-B-3-1-2-B) into the genomes of Osmancık-97 and Halilbey. The chromosome segment containing the Pi40 gene was introduced and the improved ABLs of BC generations contained limited donor derived chromosome segments in the recurrent parent background genome. Conventional molecular techniques in breeding employ the separation of PCR amplicons in agarose or polyacrylamide gels and subsequent visualization in gel documentation systems. These steps are not just laborious but more importantly, cost-ineffective (Ramkumar et al., 2015). The advent of advances in molecular breeding which include SNP genotyping technologies and automated capillary electrophoresis systems has the potential to answer these problems. Automation using capillary electrophoresis can also potentially lower the cost and time utilized during gel electrophoresis preparations. To demonstrate the applicability of these new technologies, ABLs, RPs, and DP were SNP genotyped using Illumina 6K Infinium SNP Chip. PCR amplicons from the parents and ABLs were also run in an automated capillary electrophoresis system using Fragment Analyzer. A similar strategy was performed using the Pi2 gene. Elite thermo-sensitive genic male sterile lines were introgressed with Pi2 using molecular breeding approach (Jiang et al., 2015). A whole genome SNP array similar to what is used in this study was also utilized for background genotyping. RP genome recovery computed in their study was close to the genome recoveries of ABLs produced in this study. In one of the ABLs, it was also shown that most chromosomes were Table 1 Blast disease reaction of selected advance backcross lines to blast strains using spray inoculation method and field blast screening test already converted to the RP genome except for chromosomes 1, 6, 8, and 11. Parallel to the results reported by Suh et al. (Suh et al., 2009) donor-derived segments were also prevalent in chromosome 6 and can be attributed to the foreground genotyping analysis for Pi40. These results clearly indicate the applicability of these new technologies in rice molecular breeding.
The main agronomic traits of selected ABLs were mostly similar to Osmancık-97 and Halilbey. However, some of the selected plants showed increase in plant height which may be either due to the transfer of some QTLs from the donor causing increase of plant height or the soil and other climatic conditions in the Philippines. The ABLs possess strong resistance to blast with similar phenotype of the recurrent parents. Our results indicate the advantage of using genotype and phenotype selections in early backcross generations such as BC3 or BC4 to produce ABLs having desirable agronomic traits and resistance to blast. Breeding lines were also selected with BPH resistance gene, Bph18, at early backcross generation (BC2/BC3) using marker-assisted backcross breeding without linkage drag (Suh et al., 2011).
The incorporation of a novel broad-spectrum blast resistance gene, Pi40 into elite rice cultivars is considered as an effective, economical and environment friendly breeding strategy to control the disease for stable rice production. This is the first study on the introduction of a new blast resistance gene, Pi40, into the elite japonica cultivars, Osmancık-97 and Halilbey, using DNA marker-assisted backcross breeding methods. Based on agronomic traits analysis, our results showed that advanced backcross progenies had desirable phenotypes of the recurrent parents, Osmancık-97 and Halilbey. Foreground selection using the Pi40 gene specific markers could isolate early generation selected backcross progeny lines with strong resistance to blast disease. Therefore, the transfer of Pi40 gene into Turkish cultivars is an effective strategy to improve the elite cultivars of Turkey. Further studies on hotspot locations screening of selected backcross lines will identify promising genotypes with blast resistance to develop new cultivars for Turkey.
3 Conclusion
Revealing the presence of the durable broad-spectrum resistance gene, Pi40, for blast in elite Turkish cultivars showed that marker-assisted backcrossing is an effective tool to develop and improve the cultivars, Osmancik-97 and Halilbey. Advanced genomics technology such as 6K SNP genotyping and automated capillary electrophoresis analysis of the promising lines were used to carry out genotyping of selected breeding lines possessing blast resistance, high yield potential and desirable grain quality. The Pi40 blast resistance gene will enhance rice production and productivity in the temperate regions of the world and will be highly useful in other breeding programs.
4 Materials and Methods
4.1 Plant materials
Two elite cultivars of Turkey, Osmancık-97 and Halilbey, were used as RP, and IR83260-1-1-1-5-B-3-1-2-B carrying the Pi40 gene was used as DP. Eleven ABLs of BC3F5.6 and BC4F5.6 generations were also used in this study. The development of F1 and production of early backcross generation progenies (Figure 3) were conducted in Rural Development Administration (RDA) in South Korea in 2011 and the rest of the crosses and advancement of the progenies were continued at the Plant Breeding (PB) Division of the International Rice Research Institute (IRRI), Los Baños, Laguna, Philippines. The seeds of Turkish cultivars were obtained from Trakya Agricultural Research Institute (TARI), Edirne, Turkey. Seeds of the varieties CO39, IR42, IR50, IR72, IR65482-4-136-2-2 (Pi40) and Lijiangxintuanheigu (LTH) used as checks and spreader rows in the blast nursery test were obtained from PB division of IRRI.
|
Figure 3 The development of F1 and production of early backcross generation progenies. RP= Recurrent Parent; BC=Backcross; a=selfing, foreground analysis, and phenotype selection, b=SNP genotyping |
.png)
4.2 Molecular Analysis
4.2.1 DNA extraction
Young leaves collected from 14 day-old seedlings were lyophilized in Alpha 1-4 LD-2 Freeze dryer (Martin Christ, Germany) for 72 hours at -80ºC. Leaf samples were finely cut and ground in Geno-Grinder 2010 (SPEX Sample Prep, USA) at 900 rpm for 5 minutes. A modified cetyltrimethylammonium bromide (CTAB) method (Murray and Thompson, 1980) was adapted for isolation of high-quality DNA. Concentration of DNA was quantified using the Nano Drop ND-1000 (Thermo Scientific, USA). Sample preparation, DNA extraction and quantification were performed at the Novel Gene Resources (NGR) Laboratory of PB, IRRI.
4.2.2 Foreground Analysis
The DNA marker 9871.T7E2b linked to the Pi40 gene with the following primer sequences: forward (5’-CAA CAA ACG GGT CGA CAA AGG-3’) and reverse (5’-CCC CCA GGT CGT GAT ACC TTC-3’), was used for foreground analysis (Jeung et al., 2007). The Pi40 gene-specific allele amplified 642 bp band. PCR profile is as follows: initial denaturation at 95ºC for 5 minutes; 35 cycles of the following steps: denaturation at 95ºC for 1 minute, primer annealing at 65ºC for 1 minute, extension at 72ºC for 2 minutes; final extension at 72ºC for 10 minutes and storage at 4ºC. PCR cocktail includes 2 µl of 20 ng/µl template DNA, 0.3µl of 10 pmol forward and reverse primer, 2 µl of 10× PCR buffer and 1mM dNTPs respectively, 7.4 µl of PCR-grade water and 1µl Taq DNA polymerase. Undigested PCR amplicons of 642 bp were first visualized using agarose gel electrophoresis and remaining PCR products (10 µl) were used for restriction enzyme digestion using MluC1 (New England Biolabs, USA) following manufacturer’s protocol.
4.2.3 Application in high throughput genotyping
To demonstrate the application of the Pi40 gene-specific marker in high throughput technologies, PCR amplicons of the RPs, DP, and eleven ABLs were run and analyzed using Fragment Analyzer (Advanced Analytical Inc., USA). Fragment analyzer is an automated capillary electrophoresis system that relies on fluorescence excitation and constant imaging which gives on-line detection of separation at a resolution of 2 bp. The experiment was performed using manufacturer’s instructions.
4.2.4 Background analysis
To check for background genome recovery of the 11 selected ABLs, SNP genotyping was carried out along with RP and DP using high-density genome-wide genotyping chip. The Illumina 6K Infinium SNP Chip was designed at Cornell University from which SNPs were selected to be informative within and in between O. sativa subgroups and between O. sativa and O. rufipogon (Thomson, 2014). Infinium platform detects SNP alleles by adding a fluorescence-labeled allele-specific nucleotide via single-base extension and subsequent detection of the fluorescence color (Steemers et al., 2006). SNP genotyping was carried out following the manufacturer’s protocol and chips were scanned in BeadArray reader (Illumina Inc., USA). SNP genotyping data were analyzed using GenomeStudio V2011.1 (Illumina Inc., USA). Graphical genotype of ABLs was generated using Phenogram (www.ritchielab.psu.edu).
4.3 Evaluation for blast resistance
4.3.1 Blast nursery test
Screening for blast disease resistance/susceptibility of selected ABLs was conducted at IRRI blast nursery under UBN test following the method of Suh et al. (2009). Forty to fifty seedlings of test materials were planted in two replications. LTH and CO39 were used as susceptible (S) checks while IR72 and IR65482-4-136-2-2 (Pi40) were used as resistant (R) checks during the course of evaluation. Spreader rows with a mixture of cultivars IR72, IR36, CO39, IR50 and IR42 were set up around each replicate to maintain diverse pathogen population. Scoring of the test materials was carried out 14 days after infection. Disease reaction was scored using the 0-9 scale of the standard evaluation system (SES) of IRRI (IRRI, 1996) and percent DLA.
4.3.2 Blast spray inoculation
Artificial spray inoculation of selected ABLs was carried out at the glasshouse facility of IRRI following the method described by Jeung et al. (2007) and using 21 Philippine virulent blast strains namely: M36-1-3-10-10, JMB840610, Ca23-49, V850256, V86010, M39-1-3-B-1, M39-1-2-21-2, Ca89, BN209, BN111, 43, M101-1-2-9-1, 9482-01-03, IK81-3, JMB8401, Ca41, IK81-25, P06-06, B90002, M64-1-3-9-1 and Pi9-G7-2K-1. In brief, 21-day-old seedlings were inoculated with 1.5 × 105 spores/ml blast isolate suspension. Seedlings were spray inoculated with 20 mL spore suspension and were kept in a greenhouse with 12/12 h (day/night) photoperiod at 90% relative humidity for seven days. Scoring was carried out following the method described by Japan International Research Center for Agricultural Sciences (Hayashi and Fukuta, 2009).
4.4 Agronomic and grain quality evaluation of selected ABLs
Materials were seeded in a seed box and twenty one day old seedlings were transplanted at the Experiment Station of IRRI. Recurrent and donor parents were planted along with ABLs in 5-rows with 20cm × 20cm spacing using a complete randomized design during 2014 wet season (WS) and 2015 dry season (DS). Major agronomic traits such as PH, TN, days to heading (DTH), FLL, 100 GW, PL, GN, F during 2014WS and 2015DS were collected following standard evaluation system of IRRI. Grain quality was estimated for AC, GT, GC and PV. AC is determined by colorimetric assay of the amylase-iodine complex (Juliano, 1971). GT category was estimated by determining the alkali spreading value. The degree of spreading of 6 head rice soaked in 10 mL of 1.7% KOH after 23 hours at room temperature (25 ± 5oC) was measured and a score of 1-7 is given, depending on the intensity of swelling and spreading, 1 being no reaction and 7 being mostly dissolved. Alkali spreading value of 1-2 corresponds to high GT, 3 for intermediate high, 4-5 for intermediate, and 6-7 for low GT (Little et al., 1958). Gel consistency was measured following the protocol of Cagampang (1971). In brief, the consistency of a cold 4.4% milled rice paste in 0.2 M KOH was measured by the length of the cold gel in the culture tube held horizontally for 30 min to 1 hour. PV was measured by quantification of total nitogren content by Kjeldahl digestion method. The ammonia produced in the reaction was determined colorimetrically as indo-phenol blue (IRRI, 2005, http://asl.irri.org/lims/index.jsp?page=about&feature=plantanalysis#nkm).
4.5 Statistical analysis
A one-way analysis of variance and comparison of means using Tukey’s honestly significant difference test at 95% confidence interval (p<0.05) were carried out. Agronomic data were processed and statistically analyzed using Statistical Tool for Agricultural Research software v2.0.1, a free application tool created using Eclipse Rich Client Platform and R language developed at IRRI (IRRI, 2013).
Authors’ contributions
KKJ conceived the study and wrote the manuscript. NB and HS participated in experimental design and evaluated the ABLs for blast resistance in Turkey. SMK developed the early generation progenies. MMDV advanced the progenies, carried out foreground genotyping analysis, conducted blast screening experiments, gathered agronomic data, and drafted the manuscript. RBV carried out background genotyping, statistical analysis, and drafted the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We like to thank the General Directorate of Agricultural Research and Policies, (GDAR), Ministry of Food, Agriculture and Livestock, Republic Of Turkey for funding this project (Grant # DRPC 2010-69). We thank Mr. Patricio Carandang for his technical assistance during the progress of this project. We like to extend our appreciation to Ms. Sherry Lou Hechanova, assistant scientist, PB, IRRI for her assistance during the preparation of this manuscript.
Beşer N., and Sürek H., 2012, Rice Food Security and Production in Turkey, In: Jena K.K., Hardy B. (eds.), Advances in Temperate Rice Research, International Rice Research Institute, Los Baños, Philippines, pp.89-96
Bonman J.M., and Mackill D.J., 1988, Durable resistance to rice blast, Oryza, 25: 103-110
Bonman J.M., Khush G.S., and Nelson R.J., 1992, Breeding for resistance to pests, Annu. Rev. Phytopathol., 30: 507-528
http://dx.doi.org/10.1146/annurev.py.30.090192.002451
Cagampang G.B., 1973, A gel consistency test for eating quality of rice, Journal of the Science of Food & Agriculture, 24(12): 1589-1594
http://dx.doi.org/10.1002/jsfa.2740241214 PMid:4771843
De Datta S.K., 1981, Principles and Practices of Rice Production, International Rice Research Institute, Los Baños, Philippines, pp.2-3
Fu C., Wul T., Liu W., Wang F., Li J., Zhu X., Huang H., Liu Z. R., Liao Y., Zhu M., Chen J., and Huang Y., 2012, Genetic improvement of resistance to blast and bacterial blight of the elite Rongfeng B in hybrid rice (Oryza sativa L.) using marker-assisted selection, African Journal of Biotechnology, 11(67): 13104-13124
http://dx.doi.org/10.5897/AJB12.1465
Fukuta Y., Vera-Cruz C.M., and Kobayashi N., 2009, Development and characterization of blast resistance using differential varieties in rice, JIRCAS Working Report No.63, Tsukuba, Japan, pp. iii + 122 pp
Hayashi N., and Fukuta Y., 2009, JIRCAS Research Project: Blast Research Network for Stable Rice Production, JIRCAS Working Report No. 63, Tsukuba, Japan, pp.123
IRRI, 1996, Standard Evaluation System for Rice, 4th ed. International Rice Research Institute, Los Baños, Philippines, pp.52
IRRI, 2013, Statistical Tool for Agricultural Research (STAR). Plant Breeding, Genetics, and Biotechnology Division, International Rice Research Institute, Los Baños, Philippines
Jena K.K., and Kim S.M., 2010, Current status of brown planthopper (BPH) resistance and genetics, Rice, 3(2-3): 161-171
http://dx.doi.org/10.1007/s12284-010-9050-y
Jena K.K., and Mackill D.J., 2008, Molecular markers and their use in marker-assisted selection in Rice, Crop Science, 48(4): 1266-1276
http://dx.doi.org/10.2135/cropsci2008.02.0082
Jeung J.U., Kim B.R., Cho Y.C., Han S.S., Moon H.P., Lee Y.T., and Jena K.K., 2007, A novel gene, Pi40(t), linked to the DNA markers derived from NBS-LRR motifs confers broad spectrum of blast resistance in rice, Theoretical & Applied Genetics, 115(8): 1163-1177
http://dx.doi.org/10.1007/s00122-007-0642-x PMid:17909744
Jiang J., Mou T., Yu H., and Zhou F., 2015, Molecular breeding of thermo-sensitive genic male sterile (TGMS) lines of ricefor blast resistance using Pi2 gene, Rice, 8(1): DOI 10.1186/s12284-015-0048-3
http://dx.doi.org/10.1186/s12284-015-0048-3
Juliano B.O., 1971, A Simplified assay for milled-rice amylase, Cereal Science Today, 16(10): 334-360
Khush G.S., 2005, What it will take to feed 5.0 billion rice consumers in 2030, Plant Molecular Biology, 59(1): 1-6
http://dx.doi.org/10.1007/s11103-005-2159-5 PMid:16217597
Khush G.S., and Jena K.K., 2009, Current status and future prospects for research on blast resistance in rice (Oryza sativa L.), In: Wang G.L. and Valent B. (eds.) Advances in genetics, genomics and control of rice blast, Springer Science, Netherlands, pp.1-10
http://dx.doi.org/10.1007/978-1-4020-9500-9_1 PMid:19265486
Little R.R., Hilder G.B., and Dawson E.H., 1958, Differential Effect of dilute alkali on 25 varieties of milled white rice, Cereal Chem., 35: 111-126
Manners G., 2013, Rice booms in Turkey, Rice Today, 12: 26-29
Miah G., Rafii M.Y., Ismail M.R., Puteh A.B., Rahim H.A., Asfaliza R., and Latif M.A., 2013, Blast resistance in rice: a review of conventional breeding to molecular approaches, Molecular Biology Reports, 40(3): 2369-2388
http://dx.doi.org/10.1007/s11033-012-2318-0 PMid:23184051
Murray M.G., and Thompson W.F., 1980, Rapid isolation of high molecular-weight plant DNA, Nucleic Acids Research, 8(19): 4321-4325
http://dx.doi.org/10.1093/nar/8.19.4321 PMid:7433111 PMCid:PMC324241
Neeraja C.N., Rodriguez R.M., Pamplona A., Heuer S., Collard B.C.Y., Septiningsih E.M., Vergara G., Sanchez D., Xu K., Ismail A.M., and Mackill D.J., 2007, A marker-assisted backcross approach for developing submergence-tolerant rice cultivars, Theoretical & Applied Genetics, 115(6): 767-776
http://dx.doi.org/10.1007/s00122-007-0607-0 PMid:17657470
Normile D., 2008, Reinventing rice to feed the world, Science, 321(5887): 330-333
http://dx.doi.org/10.1126/science.321.5887.330 PMid:18635770
Ou S.H., 1980, Pathogen variability and host resistance in rice blast disease, Annual Review of Phytopathology, 18(4): 167-187
http://dx.doi.org/10.1146/annurev.py.18.090180.001123
Ramkumar G., Prahalada G.D., Hechanova S.L., Vinarao R., and Jena K.K., 2015, Development and validation of SNP-based functional codominant markers for two major disease resistance genes in rice (O. sativa L.), Molecular Breeding, 35(6): 1-11
http://dx.doi.org/10.1007/s11032-015-0323-4
Sharma T.R., Rai A.K., Gupta S.K., Vijayan J., Devanna B.N., and Ray S., 2012, Rice Blast Management Through Host-Plant Resistance: Retrospect and Prospects, Agricultural Research ., 1(1): 37-52
http://dx.doi.org/10.1007/s40003-011-0003-5
Steemers F.J., Chang W., Lee G., Barker D.L., Shen R., and Gunderson K.L., 2006, Whole-genome genotyping with the single-base extension assay, Nature methods, 3(1): 31-33
http://dx.doi.org/10.1038/nmeth842 PMid:16369550
Suh J.P., Roh J.H., Cho Y.C., Han S.S., Kim Y.G., and Jena K.K., 2009, The Pi40 Gene for durable resistance to rice blast and molecular analysis of Pi40-advanced backcross breeding lines, Phytopathology, 99(3): 243-250
http://dx.doi.org/10.1094/PHYTO-99-3-0243 PMid:19203276
Suh J.P., Yang S.J., Jeung J.U., Pamplona A., Kim J.J., Lee J.H., Hong H.C., Yang C.I., Kim Y.G., and Jena K.K., 2011, Development of elite breeding lines conferring Bph18 gene-derived resistance to brown planthopper (BPH) by marker-assisted selection and genome-wide background analysis in japonica rice (Oryza sativa L.), Field Crops Research, 120(2): 215-222
http://dx.doi.org/10.1016/j.fcr.2010.10.004
Thomson M.J., 2014, High-throughput SNP genotyping to accelerate crop improvement,Plant Breeding and Biotechnology,2(3): 195-212
http://dx.doi.org/10.9787/PBB.2014.2.3.195
Zeigler R.S., Thome J., Nelson J., Levy M., and Correa-Victoria F.J., 1994, Lineage exclusion: A proposal for linking blast population analysis to resistance breeding, In: Rice blast disease, CAB International, Wallingford, U.K., pp.267-292



. PDF(1096KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Necmi Beser
. Marilyn M. Del Valle
. Suk-Man Kim
. Ricky B. Vinarao
. Halil Surek
. Kshirod K. Jena
Related articles
. Blast
. Marker-assisted backcrossing
. Pi40
. Resistance, Rice
Tools
. Email to a friend
. Post a comment