Research Report
The CDS Variance Analysis of OsBADH2 Gene and Aroma Appraisal of 7 Guizhou HE (Oryza sativa) 
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2018, Vol. 9, No. 12 doi: 10.5376/mpb.2018.09.0012
Received: 22 Nov., 2018 Accepted: 10 Dec., 2018 Published: 30 Dec., 2018
Wang L.L., Peng Q., Li J.L., Zhang D.S., Wu J.Q., Jiang X., and Zhu S.S., 2018, The CDS variance analysis of OsBADH2 gene and aroma appraisal of 7 Guizhou HE (Oryza sativa), Molecular Plant Breeding, 9(12): 87-91 (doi: 10.5376/mpb.2018.09.0012)
Dysfunction of OsBADH2 gene in rice is the internal cause of rice to have fragrance. This study applied physicochemical methods to identify the fragrance traits of 7 Guizhou HE (Oryza sativa) and 2 control varieties. It was found that RongHe and 2 control varieties were fragrant rice, and the other 6 varieties were all non-fragrant rice. To analyze the internal relationship between fragrant production in Guizhou HE and OsBADH2 gene, the study obtained the CDS sequences of OsBADH2 gene in 9 tested materials, and conducted alignment analysis of these sequences by Megalign software through TA-cloning, sequencing analysis and sequence assembly. The results showed that the OsBADH2 gene CDS of 4 Guizhou HE (BaixiangHe 2, BanpoHe, LipingHe 1 and LipingHe 2) were normal. The CDS differences of RongHe and 2 control varieties were both 8 bp deletion and 3 SNPs in exon 7, and 2 new CDS differences were found among which BaixiangHe 1 was 1 bp deletion and 1 SNP, and BianHe was 2 bp deletion and 2 SNPs. These results might provide the theoretical basis for variety protection and breeding improvement of Guizhou HE (Oryza sativa).
Background
Fragrance is an important quality trait of high-quality rice. With the continuous improvement of people’s living standards, consumers’ demand for high-quality rice is increasing. The rice with excellent quality and rich flavor has become popular in the market. Therefore, the mechanism research of rice flavor production and the breeding of aromatic rice varieties have great market value and practical significance.
As early as the 1970s, scholars began to study the volatile components of rice fragrance (Yajima et al., 1979). Based on years of research, it has been confirmed that 2-acetyl-1-pyrrolin (2AP) is the main fragrance component of jasmine rice (Grimm et al., 200l). With the rise of biotechnology and the innovation of research methods, rice fragrance gene research has made great progress. Bradbury et al. (2005) confirmed that rice fragrance was controlled by the OsBADH2 gene (betaine aldehyde dehydrogenase). The OsBADH2 gene encodes betaine aldehyde dehydrogenase, and the 8 base pair deletion and 3 base substitution of the 7th exon cause the loss of OsBADH2 protein function, and the substrate 2AP cannot be degraded and constantly accumulated, eventually leading to rice fragrance (Bradbury et al., 2005). In addition, Chen et al. (2008) also found an allele of the OsBADH2 gene, and the 7 bp deletion of the second exon of the allele also caused rice to produce fragrance. The isolation and cloning of the fragrance genes opened up new methods and ideas for the study of rice fragrance traits. Xu et al. (2011) designed functional molecular markers based on the difference between the 7th and 2nd exon sequences of OsBADH2 gene, and carried out molecular detection on 24 fragrant rice varieties. Jiang et al. (2015) identified the mutant sequence types of 27 fragrant rice varieties by making sequencing analysis on the 7th and 2nd exons of the OsBADH2 gene.
Guizhou HE is a kind of local rice resource that can adapt to special ecological environment. It has many excellent traits and it is an important biological genetic material (Ruan, 2001). In this study, two fragrant rice varieties (Yuefeng B and Dalixiang) were used as control to characterize the fragrance traits of seven Guizhou HE, and the CDS sequences of OsBADH2 gene were cloned and sequenced to explore the intrinsic relationship between the difference of CDS sequence and the fragrance of Guizhou HE rice and provide a theoretical basis for the fragrance variety protection and breeding improvement of Guizhou HE.
1 Results and Analysis
1.1 Physicochemical identification of fragrance traits
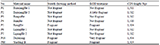
The fragrance characteristics of 9 materials (7 Guizhou HE and 2 fragrant rice controls) were identified by mouth chewing method and KOH treatment method. The identification results of these two methods were basically the same: 6 pieces of Guizhou HE materials (Baixiang HE 1, Baixiang HE 2, Banpo HE, Bian HE, Liping HE 1 and Liping HE 2) had no fragrance and were non-fragrant rice; Rong HE and the controls (Dalixiang and Yuefeng B) had fragrance and were fragrant rice (Table 1).
|
Table 1 The identification results of fragrance traits in 9 tested materials |
1.2 TA cloning for OsBADH2 gene CDS sequence
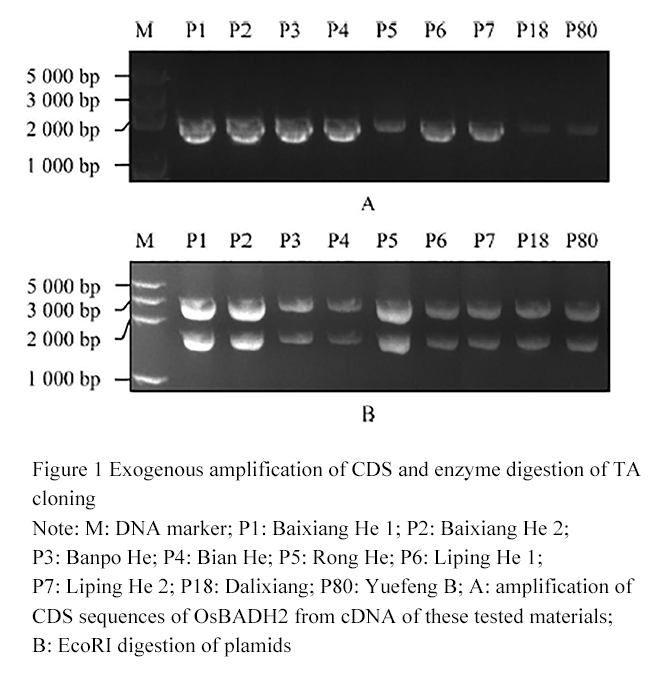
To explore whether the fragrance of the test material was caused by the OsBADH2 gene sequence difference, the leaf tissue RNA of 9 materials was extracted; the corresponding cDNA template was obtained by reversed transcription, and the OsBADH2 gene CDS were amplified with the specific primers FGR-F and FGR-R, the CDS sequences of 9 materials were amplified successfully (Figure 1A). The TA cloning of the OsBADH2 gene CDS sequences were detected by EcoRI. The exogenous DNA bands and T3 vector bands were cut out (Figure 1B), indicating that 9 CDS sequences were successfully constructed to TA cloning vectors.
|
Figure 1 Exogenous amplification of CDS and enzyme digestion of TA cloning Note: M: DNA marker; P1: Baixiang He 1; P2: Baixiang He 2; P3: Banpo He; P4: Bian He; P5: Rong He; P6: Liping He 1; P7: Liping He 2; P18: Dalixiang; P80: Yuefeng B; A: amplification of CDS sequences of OsBADH2 from cDNA of these tested materials; B: EcoRI digestion of plamids |
1.3 Sequencing CDS sequences of OsBADH2 gene
The CDS sequences of 9 materials were sequenced by SeqMan and Sequencher software. The CDS sequences of 4 materials (Banpo HE, Baixiang HE 2, Liping HE 1 and Liping HE 2) were found to be 1,512 bp in length. The length of those was the same as the known non-fragrant rice sequence. The length of the CDS sequence of Baixiang HE 1 is 1,511 bp, that of Bian HE is 1,510 bp, and the CDS sequence length is 1,504 bp for the other 3 materials (Rong HE, Dalixiang and Yuefeng B). They have the same length with the known fragrant rice sequence (Table 1).
1.4 CDS sequence alignment analysis
In order to further clarify the differences in CDS sequences of OsBADH2 gene in 9 materials, the ClastalW method of Megalign software was used for sequence alignment. It was found that among the 9 materials (7 Guizhou HE and 2 fragrant rice controls), 4 CDS sequences of OsBADH2 gene in Guizhou HE materials (Baixiang HE 2, Banpo HE, Liping HE 1 and Liping HE 2) were completely consistent with the known non-fragrant rice sequence. The CDS sequences of two fragrant rice controls (Dalixiang and Yuefeng B) and Rong HE were consistent with the fragrant rice sequence OsBADH2-E7, that is, the 7th exon had a 8 bp base deletion and 3 bp base substitutions. Baixiang HE 1 and Bian HE had new base deletions and base substitutions. CDS sequence of OsBADH2 in P1 (Baixiang HE 1) material had a base deletion at +1,060, and T/C SNP occurred at +1,399 (Figure 2). The encoded protein of P1 had a frameshift mutation from 354 aa and terminated prematurely at 360 aa, which finally encoded a truncated protein of 359 aa. The CDS sequence of OsBADH2 in P4 (Bian HE) material had a GC base deletion at +207, a C/A SNP occurred at +573, and a G/A SNP occurred at 716 (Figure 2). The encoded protein has a frameshift mutation from 70 aa, and terminated at 90 aa, eventually encoding a truncated protein of 89 aa.
|
Figure 2 The variation of CDS sequences of OsBADH2 gene Note: - indicates deletion of nucleotide, * indicates nucleotide that is different from reference CDS |

2 Discussion
Rapid and accurate identification of rice fragrance traits is a premise for fragrance research and fragrant rice breeding. KOH soaking method and chewing method are traditional rice fragrance identification methods. Considering the influence of personal subjective factors (personal taste and smell on the perception of rice fragrance), the identification results have certain errors and are time-consuming and laborious. The identification results of this research also confirmed the above viewpoints. The fragrance characteristics of 9 materials (7 Guizhou HE and 2 fragrant rice controls) were identified by chewing and KOH soaking method. It was found that 2 of the 9 materials had different identification results. Baixiang HE 2 was identified as non-fragrant by chewing, while a slight fragrance by KOH soaking method. The two methods had different identification results of Dalixiang on the degree of rice fragrance as well (Table 1). With the cloning of the fragrance gene OsBADH2 and the development of biotechnology, the identification of fragrance traits has entered to the molecular level. Molecular identification of rice fragrance traits by PCR was conducted by developing molecular markers linked to the OsBADH2 gene or designing specific primers based on the OsBADH2 gene sequence differences between fragrant and non-fragrant rice (Bradbury et al., 2005; Shi et al., 2008).
The CDS of the OsBADH2 gene in non-fragrant rice was 1,512 bp in length, while that was 1,504 bp in the fragrant rice due to the 8 bp deletion in the 7th exon and 3 SNPs (Bradbury et al., 2005). In order to test whether the identification results of traditional method are consistent with the OsBADH2 CDS sequence characteristics, this research performed TA cloning and sequencing analysis of OsBADH2 gene CDS in Guizhou HE, and obtained the CDS of 9 materials. The sequence alignment showed that the CDS sequences of two control cultivars (Dalixiang and Yuefeng B) and that of Rong He were fragrant-rice sequences, and the CDS sequences of four Guizhou HE materials (Baixiang HE 2, Banpo HE, Liping HE 1 and Liping HE 2) were non-fragrant-rice sequence (Table 1; Figure 2), which were consistent with expectations. There were new variant types (base deletions and SNP locis) in the CDS sequences of Baixiang HE 1 and Bian HE (Figure 2), and the encoded proteins could lead to frameshift and premature termination. However, both chewing and KOH soaking methods identified those as non-fragrant rice, which indicated that there were other fragrance genes or fragrance substances that affected or controlled the rice fragrance.
3 Materials and Methods
3.1 Material planting
Two fragrant rice control materials (Dalixiang and Yuefeng B), and 7 Guizhou HE were collected from Liping, Rongjiang, etc. These materials were planted in the experimental rice field of rice institute of Guizhou province (Guiyang).
3.2 Mouth chewing method to identify material fragrance
The method of chewing to identify the fragrance of rice was referred by Dhulappanavar et al. (1976). Randomly select 10 dried seeds of each material for analysis. If 10 seeds were scentless, the material was deemed as non-fragrant rice; if 10 seeds had fragrance, the material was deemed as fragrant rice; if there were fragrant and non-fragrant rice among these ten seeds, the material was classified as hybrid separation.
3.3 KOH treatment to identify material fragrance
The KOH soaking method was used to identify the leaf fragrance according to the method of Sood and Siddiq (1978). Take about 2 g of mature rice leaves at the peak of the cultivating period, cut them into small pieces and put into glass bottles containing 10 mL of 1.7% KOH solution. Cover the bottle and soak for 10 min at room temperature. After opening the bottle cap, 3 people took turns to smell and confirm the presence or absence of the fragrance.
3.4 TA cloning and sequencing of OsBADH2 gene CDS sequence
Take the mature leaves which were sown 30 days before, and extract the total RNA according to the instructions of TRIzol reagent (Invitrogen company); take 5 μg of total RNA and perform reverse transcription to amplify cDNA according to NEB’s M-MULV reverse transcriptase reverse transcription kit. Design CDS amplification primers based on OsBADH2 cDNA sequence, FGR-F: CACCACTCCACACCTGACAC and FGR-R: TTACAGCTTGGAAGGGGAT. Using 20 μL PCR system: DNA 1.0 μL, dNTPs 4.0 μL, 2xBuffer 10.0 μL, KOD FX 0.2 μL, FGR-F/FGR-R 0.3 μL each, ddH2O 4.2 μL. Amplifyed by 9700 PCR instrument, reaction procedure: 94°C for 2 min; 98°C for 10 s, 55°C for 30 s, 68°C for 100 s, 34 cycles; 68°C for 7 min. After the PCR product was tailed through A-Tailing, it was transferred to Trans1-T1 competent cells by heat shock method with pEASY®-T3 Cloning Vector. After screening with AIX plate blue and white spots, the positive monoclonal was chosen to reproduce, and the plasmid DNA was extracted by alkaline lysis method, and confirmed by EcoRI digestion, and then sent to Invitrogen for sequencing, and each sequencing reaction was repeated twice.
3.5 Sequence splicing and comparison analysis
The error-prone intervals at both ends were removed, and the sequencing results were compared and spliced using SeqMan and Sequencher software to obtain the CDS sequence of OsBADH2 of each material. The OsBADH2 gene (LOC_Os08g32870) CDS sequences of fragrant rice and non-fragrant rice were used as reference sequences, and the differences in CDS sequence of OsBADH2 gene among these tested materials were analyzed by MegAlign software.
Authors’ contributions
PQ and WLL are the designers and executors of this research experiment, responsible for the paper writing and revision; LJL is responsible for molecular cloning and sequencing analysis; ZDS and JX are responsible for the physicochemical identification of fragrance material; WJQ is responsible for material planting and management in the field; ZSS is the conceiver in charge of this research, guiding experimental design and data analysis. All authors read and approved the final manuscript.
Acknowledgments
This research was co-sponsored by the 12th Five-Year Plan of National Science and Technology in Rural Areas (2014AA10A604-11), Guizhou Provincial Joint Fund Project (Qianke He LH (2015) 7074), Guizhou Provincial Major Special Qianke (2013) 6023, Guizhou Provincial Science and Technology Support Program (Qianke He support (2018) 2298), Guizhou Provincial Scientific Research Institutions Service Enterprises Action Plan (Qian Ke He Fu Enterprise (2014) 4005) and Guizhou Provincial Science and Technology Fund (Qian Ke He J (2015) 2098).
Bradbury L.M.T., Fitzgerald T.L., Henry R J, Liu X.G., and Xu M.L., 2005, The gene for fragrance in rice, Plant Biotechnol. J., 3(3): 363-370
https://doi.org/10.1111/j.1467-7652.2005.00131.x
Chen S.H., Yang Y., Shi W.W., Ji Q., He F., Zhang Z.D., Cheng Z.K., Liu X.G., and Xu M.D., 2008, Badh2, Encoding betaine aldehyde dehydrogenase, inhibits the biosynthesis of 2-acetyl-1-pyrroline, a major component in rice fragrance, Plant Cell, 20(7): 1850-1861
https://doi.org/10.1105/tpc.108.058917
Dhulappanavar C.V., 1976, Inheritance of scent in rice, Euphytica, 25(1): 659-662
Grimm C.C., Bergman C., Delgado J.T., and Bryant R., 2001, Screening for 2-acetyl-1-pyrroline in the headspace of rice using SPME/GC-MS, Agric. Food. Chem., 49(1): 245-249
Jiang D., Lu X.Y., Wang Y.C., Fan Y.L., Zhai W.X., Luo Y.H., and Xia Z.H., 2015, Sequence analysis of mutation site of badh2 in 27 varieties of fragrant rice, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 13(2): 276-280
Ruan R.C., Chen H.C., Zhang Z.X., Yang Y.S., Jin T.Y., You J.M., and Zhu Y.Q., 2001, Progress and prospects of research and utilization on genetic diversity of rice germplasm in Guizhou, Zhiuwu Yichuan Ziyuan Kexue (Journal of Plant Genetic Resources), 2(2): 56-60
Sakthivel K., Sundaram R.M., Shobha R.N., Balachandran S.M., and Neeraja C.N., 2009, Genetic and molecular basis of fragrance in rice, Biotechnol Adv., 27: 468-473
https://doi.org/10.1016/j.biotechadv.2009.04.001
Shi W.W., Yang Y., Chen S.H., and Xu M.L., 2008, Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties, Mol. Breeding, 22(2): 185-19
https://doi.org/10.1007/s11032-008-9165-7
Sood B.C., and Siddiq E.A., 1978, A rapid technique for scent determination in rice, Indian Journal of Genetics & Plant Breeding, 38(2): 268-275
Xu X.L., Zhao G.Q., and Li J.Y., 2011, Development of molecular markers used to identify two types of fragrant rice and analysis of mutation sites of BADH2 gene in 24 varieties of fragrant rice, Zhiwu Fenlei Yu Ziyuan Xuebao (Plant Diversity and Resources), 33(6): 667-673
Yajima L., Yanai T., Nakamura M., Sakakibara H., and Hayashi K., 1979, Volatile flavor components of cooked Kaorimai (scented rice, O. sativa japonica), Agric. Biol. Chem., 43: 2425-2429
https://doi.org/10.1080/00021369.1979.10863850
https://doi.org/10.1271/bbb1961.43.2425



. PDF(345KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Lili Wang
. Qiang Peng
. Jiali Li
. Dashuang Zhang
. Jianqiang Wu
. Xue Jiang
. Susong Zhu
Related articles
. Oryza sativa L.
. Fragrance
. OsBADH2
. TA-cloning
. Sequence alignment
Tools
. Email to a friend
. Post a comment