Research Article
Genome-wide Identification and Analysis of β-galactosidase (BGAL) Gene Family in Cotton 
2 Institute of Cotton Research of the Chinese Academy of Agricultural Sciences, State Key Laboratory of Cotton Biology Cotton, Anyang; 455000, P.R. China
3 Anyang Institute of Technology, Anyang, 455000, 130033, P.R. China
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2021, Vol. 12, No. 27 doi: 10.5376/mpb.2021.12.0027
Received: 15 Sep., 2021 Accepted: 25 Sep., 2021 Published: 21 Oct., 2021
Cao X.C., Zhang C.J., Gou H.Q., Wang X.Y., Qiao K.K., Ma Q.F., Zhang G.Y., and Fan S.L., 2021, Genome-wide identification and analysis of β-galactosidase (BGAL) gene family in cotton, Molecular Plant Breeding, 12(27): 1-16 (doi: 10.5376/mpb.2021.12.0027)
Plant β-galactosidases (BGALs) are the important glycosidase that hydrolyses the non-reductive terminaβ-D-galactosidase residues from glycochains, glycolipids and glycoproteins. In order to reveal the regulation mechanism of β-galactosidases in cotton development, the whole genome analysis of BGAL gene family were carried out in this study, which laid a foundation for further understanding of the function of BGAL genes in cotton pollen. A total of 153 BGAL genes were respectively identified in Gossypium hirsutum, G.barbadense, G.arboreum, G.raimondii. Phylogenetic tree analysis showed that the BGAL genes are divided into eight subgroups, with the same number of exons and genetic structure in the same subgroup. The co-evolutionary analysis showed that there were multiple gene pairs between G.hirsutum and diploid cotton. The transcriptomic data showed that some genes in G.hirsutum were specifically expressed in different tissues. For example, GhBGAL6 and GhBGAL32, are highly expressed in all tissues; GhBGAL33, GhBGAL7, GhBGAL18 and GhBGAL43 were highly expressed in stamens (anther and filament) and petals. Further qRT-PCR results showed that some genes (such as GhBGAL6, GhBGAL7 and GhBGAL17) were highly expressed in stamens, which may have a certain regulatory effect on the development of stamens. This study explored the evolution and function of the BGAL gene family in the genomes of cotton, which can provide a theoretical basis for subsequent research on BGAL genes in cotton.
Glycoside hydrolases (GH, EC3.2.1) are glycoside bond enzymes capable of hydrolyzing between two or more carbohydrates or between carbohydrate and non-carbohydrate parts (Henrissat and Bairoch, 1993). Glycosidic hydrolases are found in almost all organisms and can hydrolyze the glycosidic bonds of various carbohydrate compounds (including monoglycosides, oligosaccharides, polysaccharides, saponins, glycoproteins, etc.) by endoscopically or exscopically to produce monosaccharides, oligosaccharides, or sugar complexes. Glycoside hydrolase has undergone structural changes in the course of evolution. According to the difference in protein structure of hydrolase, it can be divided into 135 families (Glycoside hydrolase family), namely GH1-GH135 (Henrissat and Bairoch, 1993). The structural similarity of members of each family is very high, and according to the structural characteristics of the structural domain of the catalytic active center, these families can be classified into 14 clans, namely GH-A-GH-N (Ahn et al., 2007). β-galactosidase (BGAL) only exists in the glucoside hydrolase family GH1, 2, 3, 35, 42, 50 and 59, belonging to GHA. Plant β-galactosidase (EC3.2.1.23) is found only in the GH35 family, Arabidopsis thaliana (Ahn et al., 2007), Solanum Lycopersicum (Smith and Gross, 2000), Carica papaya (Lazan et al., 2004), and Oryza sativa (Tanthanuch et al., 2008) all contain BGAL family genes. This suggests that BGAL gene diversity is prevalent in plants.
The degradation of lactose, proteoglycan, glycolipid, oligosaccharide, and polysaccharide widely present in plants, animals and microorganisms are mainly caused by the hydrolysis of the terminal non-reducing β-D-galactoside residues in β-D-galactoside (Lombard et al., 2014) by β-galactosidase (BGAL). BGALs in plants are classified into two categories: I and II. Class I is composed of β-(1→4) exigent galactanase, which can act specifically on β-(1→4) galactanase in pectin to produce galactose residues. Class II specifically hydrolyzes β-(1→3)- and β-(1→6) galactose residues of arabinogalactan-proteins (AGPs) to produce monuronic acids, but has no activity for β-(1→4) galactose in pectin (Sørensen et al., 2000). Class I BGAL has a specific effect on β-(1→4) β-galactosidase residues in pectin and xyloglucan, so class I BGAL plays an important role in cell wall structure and intercellular adhesion. Studies have reported that BGAL is involved in pectin decomposition during fruit ripening, softening the cell wall of plant fruits in a variety of plants such as kiwifruit, persimmon, sweet cherry, mango and peach (Guo et al., 2018). Class II BGAL is involved in many stages of the development of other plant tissues, such as spinach leaves, mung bean seedlings, radish hypocotyls and young leaves, and the meristem region of root, cotyledon, vascular tissue, trichome and pollen of tobacco (Hrubá et al., 2005). In addition, the seeds of Tropueolum mujus L., Copaifera langsdorffii and Hymenaea courbaril were observed in cotyledon. β-galactosidase is involved in the degradation of xyloglucan (Ahn et al., 2007), and it has been reported that BGAL acts with α-xyloglucase, β-glucosidase and other enzymes to achieve the degradation of xyloglucan (Wang et al., 2018). Therefore, BGAL plays an important role in plant cell wall remodeling.
Plant pollen wall plays an important role in pollen development, and the development of pollen wall will directly affect the process of pollen development, thus affecting the fertility of plants (Tian et al., 2014). The main component of pollen wall is relatively stable sporopollenin. In the mononucleate stage of microspore development, sporopollenin derived from tapetum will cover pollen and form pollen wall structure, which can effectively protect pollen from external influence (Moctezuma et al., 2003). During pollen maturity, pollen wall components such as sporopollenin degrade normally and release pollen. The synthesis and normal degradation of sporopollenin are both complex biological processes, which are jointly regulated by many genes (Smith and Gross, 2000). β-galactosidase and other genes are important genes involved in sporopollenin degradation (Ban et al., 2018). β-galactosidase has been associated with pollen development in a variety of species, such as AtBGAL7 and AtBGAL15 genes in Arabidopsis thaliana (Hrubá et al., 2005) and OsBGAL5, OsBGAL12, OsBGAL14 and OsBGAL15 genes in Oryza sativa L. play a role in early microspore development and pollen development. In Tobacco (Rogers et al., 2001), northern hybridization showed that BGAL family genes were specifically expressed in anthers, mature pollen grains and late microspore development, indicating that they also played a role in the growth process of pollen tubes. In Chinese cabbage (Liu et al., 2013), many genes of BGAL family are specifically expressed in pollen.
Studies have shown that BGAL is not only an essential enzyme in plant growth and development, but also affects the development of pollen wall. Although the BGAL family has been studied in several species, it has not been reported in cotton. With the rapid development of sequencing technology, the genome sequences of G.hirsutum, G.arbadense, G.Raimondii have been sequenced and analyze. In this study, bioinformatics methods and bioinformation analysis tools were used to identify and analyze the BGAL family in cotton (Lu et al., 2018), and the expression pattern of GhBGAL genes were analyzed. The qRT-PCR results showed that GhBGALs was expressed in different tissues. Furthermore, the function of BGAL genes in G.hirsutum and the tissues where it plays a role were predicted, thus providing a theoretical basis for the subsequent in-depth study of the application of BGAL gene family in G.hirsutum.
1 Results and Analysis
1.1 Identification of members of the BGAL genes in cotton
A total of 153 BGAL genes were identified and renamed according to the sequence of genes on chromosomes. Among them, there were 51 G.hirsutum, named GhBGAL1~GhBGAL51. There were 54 varieties of G.barbadense, named GbBGAL1~GbBGAL54; There were 24 varieties of G.raimondii, named GrBGAL1~GrBGAL24; There were 24 G.arboreum, named GaBGAL1~GaBGAL24 (Table 1). According to the quantitative analysis of BGAL genes in different species, the number of BGAL family in four cotton species were higher than that in Arabidopsis. The number of tetraploid cotton genes were more than twice as many asdiploid cotton, which was consistent with the doubling of tetraploid cotton in BGAL genes. The number of BGAL gene family in the diploid cotton species G.arboreum and G. raimondii are the same, and the positions in chromosomes are roughly the same, indicating high homology. The physicochemical properties of the identified BGAL family showed that the length of the BGAL family proteinsranged from 335 to 891 amino acid residues (aa). The lengths of GbBGAL26, GaBGAL13 and GaBGAL23 were all greater than 1 000 aa; The molecular weights of most proteins were in the range of 70~100 kDa, with the largest molecular weight of 170.819 kDa and the smallest molecular weight of 36.612 kDa. The isoelectric points ranged from 4.90 to 9.457, and the number of exons ranged from 9 to 20. According to the prediction of signal peptides, all 139 genes had signal peptides at the n-terminal, while the other 14 genes had no signal peptides. Due to the large number of BGAL gene family, the physical and chemical properties of different cotton species were different, and the physical and chemical properties such as protein length, relative molecular weight and isoelectric point of G.arboreum were higher than those of other cotton.
.png) |
1.2 Phylogenetic analysis of BGAL protein in cotton
In order to understand the evolutionary relationship of BGAL gene family in cotton, the contiguous analysis of the amino acid sequences of BGAL family in four cotton species (G.hirsutum, G.arboreum, G.barbadense, and G. raimondii) and Arabidopsis thaliana was performed using MEGA 7.0 software, and the phylogenetic tree was constructed (Figure 1). According to our results the BGAL is divided into 4 large groups A, B, C and D. Group A was divided into five subgroups, A1 to A5, and group C was divided into C1 and C2. Group A has the largest number of family members, 88 in total. Among them, group A1 has the largest distribution, which contains 64 family members. There were only 7 BGAL gene members in both A2 and A4 groups, among which there was only one G.hirsutumgene GhBGAL6 in A2 subgroup and two G.hirsutum genes in A4 subgroup, respectively GhBGAL19 and GhBGAL45. Group B contains 31 members and has no subfamily classification. There are two subgroups in group C, consisting of 45 family members. Subgroup C1 contains only 3 members of the Arabidopsis BGAL genes. There were 40 members of the cotton BGAL genes in subgroup C2. In group D, there were 5 cotton BGAL family members and 1 Arabidopsis BGAL genes.
.png) Figure 1 phylogenetic tree of BGAL gene members |
1.3 Chromosome distribution and collinearity analysis of BGAL genes in cotton
The BGAL genes were used for chromosome localization (Figure 2). According to our results, it was found that the distribution of genes was uneven, and some chromosomes or certain regions were closely arranged. Among them, 51 GhBGALs family genes were located on 20 chromosomes, except A04, A08, A09, D04, D08 and D09. Except for A04, A08, A09, A13, D08 and D09 chromosomes, the other 20 chromosomes of 53 GbBGALs genes were distributed in G.barbadense. In addition, GbBGAL50 gene was located in scaffold (D13) fragment, and the location of BGAL family genes in G.barbadense and G.hirsutum was roughly similar. Suggesting evolutionary similarities between different species; Among the 24 BGAL genes in G.arboreum, 23 were distributed in Chr02, Chr03, Chr04, Chr05, Chr06, Chr07, Chr10, Chr11, Chr12, Chr13, and GaBGAL24 existed in scaffold (tig00008658). A total of 24 BGAL members were distributed on the other 11 chromosomes except Chr04 and Chr06.
.png) Figure 2 Chromosomal distribution of BGAL genes in cotton |
In order to understand the evolutionary relationship of BGAL family genes, collinear analysis was performed on the BGAL family genes of diploid G.arboreum, G. raimondii and tetraploid G.hirsutum (Figure 3). There were 19, 20 and 21 collinear gene pairs respectively in the subgenomes of G.arboreum and G.hirsutum, AD of G.hirsutum, and between G.hirsutum and G.raimondii. According to our research, there is a closer evolutionary relationship between G.hirsutum and G.raimondii. The above results indicated that genome rearrangement of BGAL family genes occurred in the process of polyploidy.
 .png) Figure 3 The collinearity of BGAL genes in the A genome of G.barbadense, the AD subgenome of G.hirsutum and the D genome of G.arboreum Note: The gray lines represent collinear relationships within different genomes, and the red lines represent collinear gene pairs in the BGAL genes |
1.4 Sequence alignment and structure analysis of GhBGALs genes
In order to further understand the evolution of the BGAL family of G.hirsutum, the exon-intron structure of 51 BGAL genes of G.hirsutum was studied (Figure 4), and it was found that the number of exons in BGAL genes of G.hirsutum was high. Except for 9 exons in GhBGAL16 and GhBGAL25 genes, all other genes have 10 to 20 exons (including 4 exons in 16 exons, 3 exons in 17 exons, 19 exons in 18 exons, 16 exons in 19 exons, and 1 exon in 20 exons). The number of introns in the BGAL genes is large, and the distribution of introns in most genes is very dense. Introns of A2, A5, and some of the C2 subfamily genes are scattered. Analysis results of the conserved motifs of G.hirsutum BGALs showed that motif 3, motif 4, motif 5 were found in all 7 subfamilies of G.hirsutum, indicating that these three motifs are the most conserved motifs in the BGALs genes, but the function of these conserved motifs remains to be studied. In addition, among the seven subgroups, subgroup C and subgroup A have the most motif species, but subgroup A has one more motif 6 than subgroup C, and the two subgroups A and C may be the most similar in function. The results showed that all GhBGALsgenes in the same subfamily had similar gene structure and conserved motif, which strongly supported the reliability of phylogenetic and evolutionary classification (Figure 4).
.png) Figure 4 phylogenetic tree, conserved motif and gene structure of BGAL proteins in G.hirsutum |
1.5 Analysis of promoter elements of GhBGALs
In order to understand the transcriptional regulation and potential function of GhBGALs, it is important to study the cis-acting elements in its promoter region. GhBGALs has many cis-acting elements, which can be roughly divided into three categories: A, B and C (Figure 5). A is the plant hormone response element, including auxin response element, gibberellin response element, salicylic acid response element, abscisic acid response element, methyl jasmonate response element and flavonoid response element. B is stress response element, including stress response element, drought response element, damage response element and low temperature response element; C is other responsive elements, including those related to the development of palisade mesophyll tissue, photoperiod regulatory elements, seed development, meristem development, and endosperm development. Among all GhBGALs cis-acting elements (Figure 6), the number of plant hormone-responsive elements was the largest, and almost every GhBGAL genes had one or two plant hormone-responsive elements, among which abiotic acid response elements and methyl jasmonate response elements were the largest. Among the stress response elements, drought response elements and low temperature response elements were the most numerous, and one third of GhBGALs promoters contained these two stress response elements. The number of other response elements is relatively small, generally only exist in a few genes, such as GhBGAL1, GhBGAL2, GhBGAL5, GhBGAL13, GhBGAL115 genes photoperiod regulatory elements; Seven GhBGALs promoter sequences contained expression response elements in endosperm.
.png) Figure 5 GhBGALs promoter cis-acting element Note: A: Hormone response element; B: Stress response element; C: Other response elements |
|
Figure 6 GhBGALs promoter cis-acting element |
.png)
1.6 Tissue specific expression pattern and qRT-PCR analysis of GhBGALs
In order to understand the expression of BGAL genes in G.hirsutum, 51 BGAL genes in different tissues (anther, filaments, pistil, bracts, sepals, petals, torus, roots, leaves, and stems) in G.hirsutum were analyzed based on tissue transcriptome data (Figure 7). According to the results, different genes (such as GhBGAL43 and GhBGAL38) were expressed significantly differently in the same tissues (such as anther, filaments and petals), and the same genes (such as GhBGAL42) were expressed significantly differently in different tissues (such as stamen and pistil). The expression pattern graph was clustered according to rows, and the overall analysis of genes expression differences could be divided into three parts: high expression, no expression and low expression. The expressions of 22 genes from GhBGAL46 to GhBGAL2 in all tissues showed no significant difference and were all in the state of low expression or even no expression. From GhBGAL42 to GhBGAL43 at the top of the pattern diagram, these 9 genes were highly specifically expressed in stamens (anther and filament), petals and sepals, and were less expressed in other tissues. The 20 genes from GhBGAL38 to GhBGAL37, shown at the bottom of the pattern diagram, were generally expressed at low levels in all tissues. GhBGAL6 and GhBGAL32 were highly expressed in all tissues. GhBGAL12 and GhBGAL37 were specifically expressed only in receptacle and pistil. The two genes, GhBGAL6 and GhBGAL32, were not expressed only in stems, and their expressions were low in other tissues. GhBGAL33, GhBGAL7, GhBGAL18and GhBGAL43 were highly expressed in stamens (anthers and filaments) and petals. GhBGAL22 was highly expressed in the pistil. GhBGAL20 was highly expressed in roots and stems, but low in other tissues. GhBGAL29 was highly specifically expressed in leaves. GhBGAL38 was specifically expressed in torus, root and stem, but hardly expressed in other tissues.
.png) Figure 7 GhBGALs cluster expression pattern in different tissues and organs of G.hirsutum |
In order to further understand the effect of BGAL y genes on plant tissues of G.hirsutum, 12 highly expressed BGAL genes were randomly selected in anther, filaments, pistil, bracts and sepals for qRT-PCR analysis (Figure 8). The gene GhBGAL7 is specifically expressed only in the stamens, and the expressions of GhBGAL6, GhBGAL7, GhBGAL17, GhBGAL18, GhBGAL33, GhBGAL41,GhBGAL42 and GhBGAL43 in the stamens are significantly higher than those in the pistils, bracts and sepals. These genes may regulate the development of stamens. The primers used in this study are as follows (Table 2).
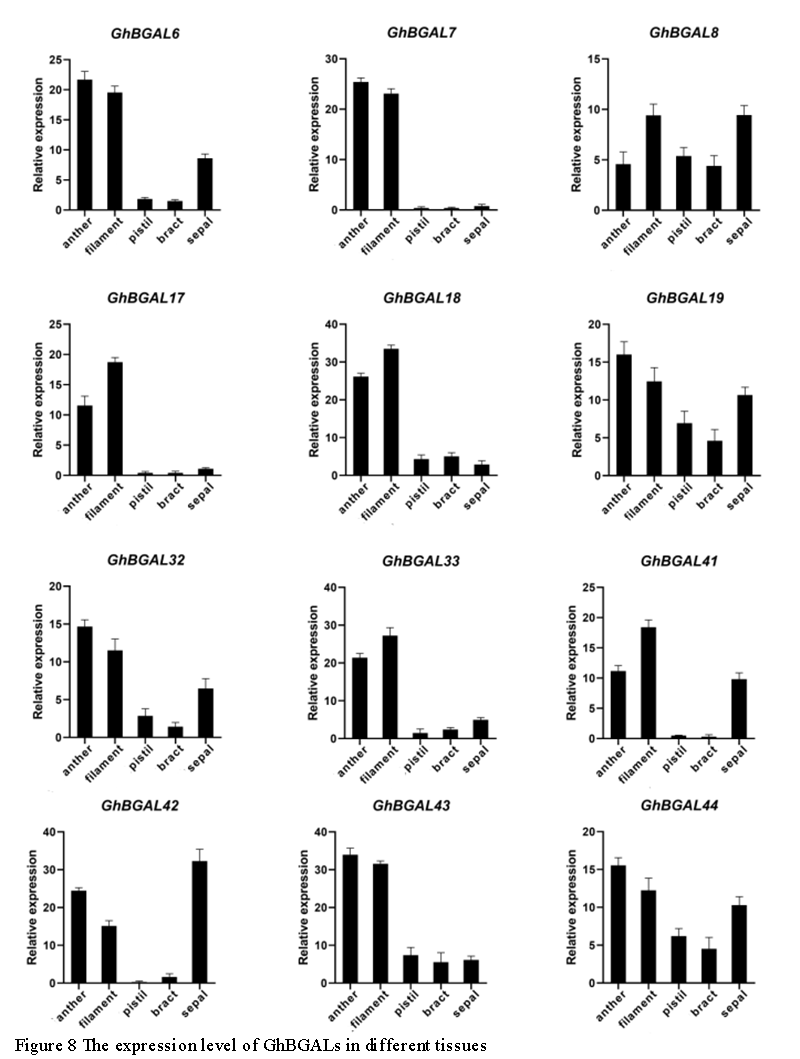 Figure 8 The expression level of GhBGALs in different tissues |
.png) |
2 Discussion
β-galactosidase can hydrolyze pectin, and has the function of softening fruit and promoting cell wall metabolism during fruit ripening. In this study, 153 BGAL genes were identified from G.hirsutum, G.barbadense, G. raimondii and G.hirsutum by bioinformatics, including 51 in G.hirsutum and 17 in Arabidopsis thaliana. The BGAL genesy has its typical sequence structure, and its sequence is GGP (LIVW)2-X(2)-Q-X-E-N-E. Multiple sequence comparison and analysis of the protein sequence of BGAL genes in cotton showed that the typical sequence of 5 genes was incomplete and part of Cys residues were missing. The other 148 genes in the cotton family all had relatively complete typical sequences, which were the sites of β-galactosidase specific binding substrates (Ahn et al., 2007). The protein sequence alignings supported the identification results of genes members.
In the phylogenetic analysis, most of the identified BGAL genes in cotton appear in pairs with high homology, and these genes may also have certain similarities in function. The 153 BGAL genes of cotton were divided into 8 subgroups, consistent with the grouping of Arabidopsis thaliana, indicating that BGAL genes of cotton have homology with Arabidopsis thaliana genes (Figure 2), but some protein sequences show differences in the evolutionary process, which may be related to subfunctionalization and natural selection of species. There is no subgroup A3 in the phylogenetic tree group because it is a bryophyte specific cluster (Ahn et al., 2007). There are noBGAL genes in the C1 subfamily, but only AtBGALs genes, which may be due to the lack of conserved protein sequences in this subfamily, and the loss of BGAL genes in the evolution of cotton. The number of genes in subgroup D is at least six, but contains BGALgenes of each species, which may be related to the conservation of genes in subgroup D. Subgroup A1, the most intensively studied among other species, encodes β-galactosidase, hydrolyzing β-(1,3)-and β-(1,4)-lactose oligosaccharide in cell walls.
Chromosome location analysis showed that the BGAL family members of G.hirsutum, G.arboreum and G.barbadense were randomly distributed on different chromosomes, while the 26 BGAL members of G.raimondii were mainly distributed on chromosomes 8, 9, 10 and 11. The number of genes in the tetraploid cotton species G.hirsutum and G.barbadense was twice that of the diploid cotton species G. raimondii and G.arboreum. It is possible that the BGAL y genes were not lost in the evolution of cross doubling in the tetraploid cotton species G.hirsutum and G.barbadense. In order to better understand the relationship between cotton species evolution, collinearity analysis was performed between different cotton species and AD subgenomes of G.hirsutum. It was found that the phenomenon of chromosome dislocation and even inversion existed in cotton, and there were coevolutionary gene pairs at some chromosomal sites, which may also have similar functions.
Analysis of conserved domain results showed that except for subgroup D, all GhBGALs genes contained motif 1, and the structures of the three subgroups A, B, and C were almost all similar, indicating that they may have similar functions. The gene expression pattern diagram could be systematically analyzed according to subgroup classification. Analysis of cis-acting elements in GhBGALs promoters reveals that almost every gene has a plant hormone responsive element. Plant hormones such as auxin and methyl jasmonate play an important role in the regulation of plant growth and development. The regulation of MeJA in the process of flower development mainly involves filament elongation, pistil development, anther development and anther dehiscing and other physiological processes. MeJA plays an important role in regulating anther dehysis in many plants, such as wheat, rice and rapeseed. Therefore, it is speculated that the response elements of plant hormones in the promoter sequence of GhBGALs may regulate the anther development of cotton.
Expression pattern analysis, GhBGAL14 and GhBGAL39 in cotton organizations almost all is in not express state, in the analysis of sequence alignment, the two gene sequences of BGAL typical incomplete, lead to can't specific to identify the role of β-galactosidase glucoside enzyme loci, cannot provide the energy for plant growth and development, not express in various organizations. β-galactosidase glucoside enzyme catalyzed by the cell walls of large and complex side chains on the metabolism of galactose and metabolism in the cell wall, the family of BGALs specific roles in plant glucan of pectin in plant cell walls, wood or Arab galactose protein degradation, in the process of mature plants, BGALsexpansion of cell wall conduction, degradation and signal molecules play an important role. BGAL family in mature fruit and seed development process of more, such as tomato and arabidopsis thaliana, a new study suggests that the yeast expression tomato TBG4 can hydrolysis of plant cell wall substrates, alkali soluble in tomato fruits during ripening stage and chelating activity of pectin mol, highest homologous genes AtBGAL4 as hydrolysis enzyme involved in the degradation of pectin, GhBGAL17 and GhBGAL42, which have the highest homology with GhBGAL17 and GhBGAL42 in G.hirsutum, are highly expressed in the reproductive organs of cotton, and these two genes may affect the development of flower organs by specifically hydrolyzing the pectin components in the cell wall or pollen wall of flowers.
Studies in recent years have found that the BGAL family influences pollen development and fertility of plants by regulating the development of pollen wall, and genes such as β-galactosidase participate in the degradation of pollen wall during the anabolic process of pollen wall. AtBGAL16 in Arabidopsis is expressed specifically in mature pollen grains, and GhBGAL32 in G.hirsutum is expressed specifically in stamens (filaments and anthers), suggesting that this gene may regulate anther development to some extent.AtBGAL7 and AtBGAL15 belong to group B genes in Arabidopsis thaliana, which, like OsBGAL5, OsBGAL12, OsBGAL14 and OsBGAL15 genes of group B in rice, play an important role in the early microspore stage of pollen development and pollen development. OsBGAL15 is highly expressed in flower organs. Studies on BGAL family transcription in fertile and sterile Chinese cabbage (Liu et al., 2013) found that BcBGAL7 and BcBGAL7 were specifically expressed in the anthers of fertile plants. In the expression pattern of G.hirsutum, GhBGAL41 in the B subgroup of GhBGALs is highly expressed in the anther (anther and filament) and petal, which is consistent with the expression in Arabidopsis thaliana, rice and Cabbage. It is suggested that GhBGAL41 may affect the development of pollen by regulating the development of pollen wall of cotton, which provides a theoretical basis for the study of male sterility of cotton.
3 Materials and Methods
3.1 Identification of BGAL family members and analysis of physical and chemical properties
Download the genomic, CDS and protein sequences of the four cotton species (Gossypium hirsutum, ZJU; Gossypium arboretum, JGI; Gossypium barbadense, ZJU; Gossypium raimondii, CRI) from Cotton Functional Genomics Database (CottonFGD) (https://cottonfgd.org/). Download biological information on other species such as Arabidopsis from the JGI database (http://www.phytozome.net). Download the configuration file of the GH35 conservative domain (PF01301) HIDDEN Markov Model (HMM) from the Pfam database (http://pfam.xfam.org). HMMER 3.0 and BLASTP were used to search for BGAL genes in the genomes of cotton and other species. Redundant genes were removed from the HMM and BLASTP results. The remaining genes were further identified by SMART (http://smart.embl-heidelberg.de/). The physicochemical properties of proteins such as amino acid length and isoelectric point (pI) of all BGAL family members of G.hirsutum, Sea G.arboreum, G.arboreum and G. raimondii were retrieved from the Cotton Functional Genomics Database (Cotton FGD) (https://cottonfgd.org/). Using cNLS Mapper (http://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi) and NetNES1.1 server (http://www.cbs.dtu.dk/services/NetNES/) to check and ratify a signal BGAL gene (NLS) and nuclear export signal (NES).
3.2 Phylogenetic tree construction of BGAL genes
The identified BGAL gene members were searched and extracted for the protein sequences of cotton and Arabidopsis by HMMER 3.0 and BLASTP, and multiple sequence alignings were performed in MEGA 7.0 software. Use online software Evolview (https://www.omicsclass.com/article/671), beautify the evolutionary tree.
3.3 Chromosome localization and coevolutionary analysis of the BGAL genes
Biological information such as location and structure of BGAL family members were extracted from the gff3 annotation files of the genome of four cotton species. MapChart2.2 software was used to analyze and map the position information of BGAL genes on chromosomes in cotton. In order to reveal the coevolutionary relationship between BGAL families among cotton species, Circos was used to construct a coevolutionary analysis diagram. According to the sequence length of the alignment covering more than 70%, the similarity of the alignment region more than 70%, the gene duplication was determined. Beautify with Adobe Illustrator CC 2019.
3.4 Gene structure and conserved motif analysis of GhBGALs
Use of MEME (http://meme.sdsc.edu/meme/) for GhBGALs conservative motif is analyzed. Use online software GSDS (http://gsds.cbi.pku.edu.cn) gene exon-contains substructure analysis GhBGALs. The analyzed sequence alignment result files, exon-intron structure files and conservative domain files were combined and visualized using TBtools.
3.5 Analysis of promoter elements of GhBGALs
In order to study the relationship between BGAL family and hormones and stress, the sequence of 1 500 bp upstream of initiation codon (ATG) in BGAL family was selected from the genome sequence of G.hirsutum. Using PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) to identify and analyze gene upstream region of cis elements, after screening using TBtools to visualize it. In order to facilitate statistical analysis, all components are classified and sorted, and histogram is made.
3.6 Tissue expression pattern analysis of GhBGALs
To understand the expression of GhBGALs in different tissues. From NCBI SRA (Sequence read archive) database (http://www.ncbi.nlm.nih.gov/sra) download G.hirsutum in different organizations (PRJNA248163) transcriptome sequencing data. Based on the characteristics of GhBGALs family, the expression levels of representative tissues such as roots, stems, leaves, anthers, filaments, pistils, bracts, sepals, petals, and torus were selected to analyze the original data. The expression levels of genes were calculated by FPKM, and the data were visualization by TBtools.
3.7 RNA isolation and quantitative reverse transcription-polymerase chain reaction (qRT-PCR) of GhBGALs
G.hirsutum 'cv CCRI24' provided by Institute of Cotton Research of the Chinese Academy of Agricultural Sciences was planted in Anyang greenhouse, sampled at full flowering stage and stored at -80℃. Total RNA was extracted using the RNAprep Pure Plant Plus kit (Tiangen, Beijing, China) according to the instructions. 2 μg total RNA was reverse transcribed into cDNA using PrimeScript first strand cDNA synthesis Kit (TaKaRa, Dalian, China), and diluted for later use. Real-time PCR was performed using SYBR premixed Ex Taq (TaKaRa, Dalian, China) in ABI 7500 system (Applied Biosystems, Foster City, CA, USA). The system consisted of 20 μL, 10 μL SYBR Green PCR mix, 0.5 μL upstream and downstream primers, 2 μL diluted cDNA, and 7 μL ddH2O. The reaction procedure of qRT-PCR was: 94℃ for 30 s; Cycle stage: 94℃ 5 s, 55℃ 15 s, 72℃ 10 s, 45 cycles; Dissolution curve stage: 94℃ 15 s, 60℃ 15 s, 95℃ 15 s, 4℃ storage. 2-△△CT method was used to calculate the relative expression level of genes using Actin as the internal reference. Each tissue and each gene expression response had 3 biological replicates and 3 technical replicates.
Authors’ contributions
CXC is the experimental design and experimental research executor of this study; CXC completed the data analysis and wrote the first draft of the paper; ZCJ, GHQ, WXY, MQF and QKK participated in experimental design and experimental result analysis; ZGY and FSL were the architects and principals of the project, guiding experimental design, data analysis, paper writing and modification. All authors read and approved the final manuscript.
Acknowledgments
This research was supported by the National Natural Science Foundation of China for Young Scholars (31701474).
Ban Q.Y.., Han Y., He Y.H..,Jin M.J., Han S.K., Suo J.T., Rao J.P., 2018, Functional characterization of persimmon β-galactosidase gene DkGAL1 in tomato reveals cell wall modification related to fruit ripening and radicle elongation, Plant Science, 274: 109-120
https://doi.org/10.1016/j.plantsci.2018.05.014
Smith D.L., and Gross K.C., 2000, A family of at least seven β-Galactosidase genes is expressed during tomato fruit development, Plant Physiology, 123(3): 1173-1183
https://doi.org/10.1104/pp.123.3.1173
Guo S.L., Song J., Zhang B.B. Jiang H., Ma R.J., Yu M.L. 2018, Genome-wide identification and expression analysis of beta-galactosidase family members during fruit softening of peach [Prunus persica (L.) Batsch], Postharvest Biology and Technology, 136: 111-123
https://doi.org/10.1016/j.postharvbio.2017.10.005
Henrissat B., and Bairoch A., 1993, New families in the classification of glycosyl hydrolases based on amino acid sequence similarities, Biochemical Journal, 293: 781-788
https://doi.org/10.1042/bj2930781
Hrubá P., Honys D., Twell D., Capková V., and Tupy J., 2005, Expression of β-galactosidase and β-xylosidase genes during microspore and pollen development, Planta, 220(6): 931-940
https://doi.org/10.1007/s00425-004-1409-0
Lazan H., Ng S.Y., Goh L.Y., and Ali Z.M., 2004, Papaya beta-galactosidase/galactanase isoforms in differential cell wall hydrolysis and fruit softening during ripening, Plant Physiology and Biochemistry, 42(11): 847-853
https://doi.org/10.1016/j.plaphy.2004.10.007
Liu J.L., Gao M.H., Lü M.L., and Cao J.S., 2013, Structure, evolution, and expression of the β-galactosidase gene family in Brassica campestris ssp. chinensis, 31(6): 1249-1260
https://doi.org/10.1007/s11105-013-0585-0
Lombard V., Golaconda R.H., Drula E., Coutinho P.M., and Henrissat B., 2014, The carbohydrate-active enzymes database (CAZy) in 2013, Nucleic Acids Research, 42: D490-495
https://doi.org/10.1093/nar/gkt1178
Lu Q., Shao F.J., and Qiu D.Y., 2018, Genome-wide analysis of gene family of lateral organ boundaries domain in Populus trichocarpa, Jiyinzuxue yu Yingyong Shengwuxue (Genomics and Applied Biology), 37(1): 313-325
Moctezuma E., Smith D.L., and Gross K.C., 2003, Antisense suppression of a β-galactosidase gene (TBG6) in tomato increases fruit cracking, J. Exp. Bot., 54(390): 2025-2033
https://doi.org/10.1093/jxb/erg214
Sørensen S.O., Pauly M., Bush M., Skjet M., Maureen M.C., Bernhardt B., Ulvskov P., 2000, Pectin engineering: modification of potato pectin by in vivo expression of an endo-1,4-β-D-galactanase, 97(13): 7639-7644
https://doi.org/10.1073/pnas.130568297
Rogers H.J., Bate N., Combe J., Sullivan J., Sweetman J., Swan C., Lonsdale D.M., and Twell D., 2001, Functional analysis of cis-regulatory elements within the promoter of the tobacco late pollen gene g10, Plant Mol. Biol., 45(5): 577-585
https://doi.org/10.1023/A:1010695226241
Tanthanuch W., Chantarangsee M., Maneesan J., Ketudat-Cairns JJJJJ., 2008, Genomic and expression analysis of glycosyl hydrolase family 35 genes from rice (Oryza sativa L.), BMC Plant Biology, 8(1): 84-80
https://doi.org/10.1186/1471-2229-8-84
Tian A.M., Liu J.L., and Cao J.S., 2014, Beta galactosidase in plants, Zhongguo Xibao Shengwuxue Xuebao (Chinese Journal of Cell Biology), 36(5): 703-707
Wang P., Li H., Jian Y., Lv Y.Z., and Gai J.T., 2018, Gene mining and evolutionary analysis of xylosyltransferase gene family in solanaceae, Jiyinzuxue yu Yingyong Shengwuxue (Genomics and Applied Biology), 37(1): 332-338
Ahn Y.O., Zheng M.Y., Bevan D.R., Esen A., Shiu S.H., Benson J., Peng H.P., Miller J.T., Cheng C.L., Poulton J.E., Shih M.C., 2007, Functional genomic analysis of Arabidopsis thalianaglycoside hydrolase family 35, Science Direct, 68(11): 1510-1520



. PDF(2224KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Xiaocong Cao
. Chaojun Zhang
. Haoqi Gou
. Xiaoyan Wang
. Kaikai Qiao
. Qifeng Ma
. Guiyin Zhang
. Shuili Fan
Related articles
. β-galactosidases
. Bioinformatics
. Cotton
. Gene family
Tools
. Email to a friend
. Post a comment