Genetics of Partial Resistance to Leaf Rust (Puccinia triticina Erikss. & Henn.) in Bread Wheat (Triticum aestivum L. em Thell) 
2 Directorate of Wheat Research (DWR) Regional Station, Flowerdale, Shimla 171002, India
3 Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi 221005, India
 Author
Author  Correspondence author
Correspondence author
Plant Gene and Trait, 2015, Vol. 6, No. 9 doi: 10.5376/pgt.2015.06.0009
Received: 05 Jul., 2015 Accepted: 27 Aug., 2015 Published: 24 Dec., 2015
The genetics of partial resistance to leaf rust (Puccinia triticina Erikss. & Henn.) in bread wheat (Triticum aestivum L. em Thell) was investigated in a nine-parent diallel (excluding reciprocals) using area under disease progress curve (AUDPC) and final disease severity (FDS) in the F1 and F2 generations. Test material consisted of slow rusting and susceptible lines. The Vr-Wr graphs exhibited partial dominance for FDS and complete dominance for AUDPC (F1 and F2generations). Parents CPAN1796, HD2329, NW1014 and MACS2496 had preponderance of minor genes for partial resistance, whereas, susceptible parents SONALIKA and HS1138-6-4 showed absence of minor genes for both AUDPC and FDS (F1 and F2 generations). Variations due to additive and dominance genetic variances were significant for both AUDPC and FDS (F1 and F2 generations). The estimate of 1-3 minor gene(s) in the slow rusting parents may be responsible for race non-specific adult plant resistance. Since narrow sense heritability for both AUDPC and FDS, was moderate in both F1 and F2, success in selecting desirable lines with partial resistance to leaf rust in early segregating generations can easily be achieved. The parents CPAN1796, NW1014, HD2329, PBW373 and RAJ3077 with low AUDPC and FDS having partial resistance either due to Lr34 and/ or minor genes were identified as good general combiners. The parents identified in the study can be used in breeding program for developing high yielding cultivars with durable leaf rust resistance.
Leaf rust, caused by Puccinia triticina Erikss. & Henn. (Syn=P. recondita Roberge ex Desmaz. f. sp. tritici) is the most widely distributed and probably causes more yield losses than the other wheat rusts (Roelfs et al., 1992; Samborski, 1984). Incorporation of genetic resistance has been used world over and found to be more economical and environment friendly method of controlling rusts. However, resistance based on major genes has not proved reliable as new races of rust pathogen keep evolving leading to boom and bust cycle. Many such examples are documented in literature including the recent emergence of Ug99 in East Africa. Partial resistance/ slow rusting conceptualized first by Caldwell (1968) appeared to be a better alternative, since it was due to number of minor genes (Knott, 1982). Singh et al. (2011) suggested use of partial resistance as the pathogen is subjected to least pressure with this type of resistance, retarding evolution of new virulences. Genetic studies based on the area under disease progress curve have often shown that race non-specific adult plant resistance to leaf rust is under oligogenic or polygenic control having additive effects with relatively high heritability (Das et al., 1993; Kuhn et al., 1980; Shaner et al., 1997). The first identified slow rusting leaf rust resistance gene Lr34 located on chromosome arm 7DS has maintained its moderate effectiveness for over 60 years (Krattinger et al., 2009). In recent years new slow rusting resistance genes namely Lr46, Lr67 and LrP (in CIMMYT wheat ‘Parula’) have been identified and located on chromosomes 1BL (William et al., 2003), 4DL (Herrera-Foessel et al., 2011; Hiebert et al., 2010) and 7BL (Singh et al., 2011), respectively. These slow rusting genes have been shown to be associated with leaf tip necrosis (Singh et al., 2011). Singh et al. (2001) selected genotypes with 2-3 minor additive genes along with Lr34 to achieve higher levels of race non-specific resistance. Substantial economic benefits have been associated with the development of such race non-specific resistance to leaf rust in many wheat producing countries, especially in areas where farmers change cultivars slowly (Smale et al., 1998). Minor genes, are generally present with major genes in most of the modern cultivars, so to study minor genes in isolation is difficult. Since major genes mask the effect of minor genes, it is imperative that we nullify the confounding effect of such genes by inoculating the target plants with appropriate races which are virulent on major genes. Present study attempts to understand the genetics of partial resistance to leaf rust through i) multipathotype test on selected wheat accessions for postulating genes and choosing appropriate pathotypes virulent on those genes ii) estimation of number of genes that confer partial resistance iii) characterization of gene action and iv) determination of general and specific combinations among selected wheat genotypes having enhanced levels of durable leaf rust resistance.
1 Results and Analysis
1.1 Postulation of major genes
On the basis of the seedling reaction test, specific leaf rust races were selected that were virulent on the major gene(s), Lr1, Lr10, Lr13, Lr23 and Lr26, postulated in the parents and are presented in Table 1. The presence of Lr34 in parents (HD2329, PBW373 and VL738) was detected with the expression of post flowering leaf tip necrosis. The seedling response to most virulent race 77-5 was a compatible reaction (susceptible) thus indicating nullification of the effect of any major gene present in parents. The adult plant reaction of parents, however, were low to moderate for AUDPC and FDS (Table 1) against mixture of different races including race 77-5 indicating the presence of minor genes for slow rusting in parents.
|
|
.png)
1.2 Test of assumption for the additive-dominance model
The validity of additive-dominance model was satisfied as the uniformity test (t2) for AUDPC and FDS showed non-significance in the F2, indicating homogeneity of Vr and Wr values. The regression coefficients ‘b’ for both AUDPC and FDS (F1 and F2) were significantly different from zero, indicating linear relationship between Wr and Vr values. However, the presence of epistasis was demonstrated in the F1 (AUDPC and FDS) and F2 (AUDPC) as ‘b’ deviated significantly from unity.
1.3 Graphic analysis
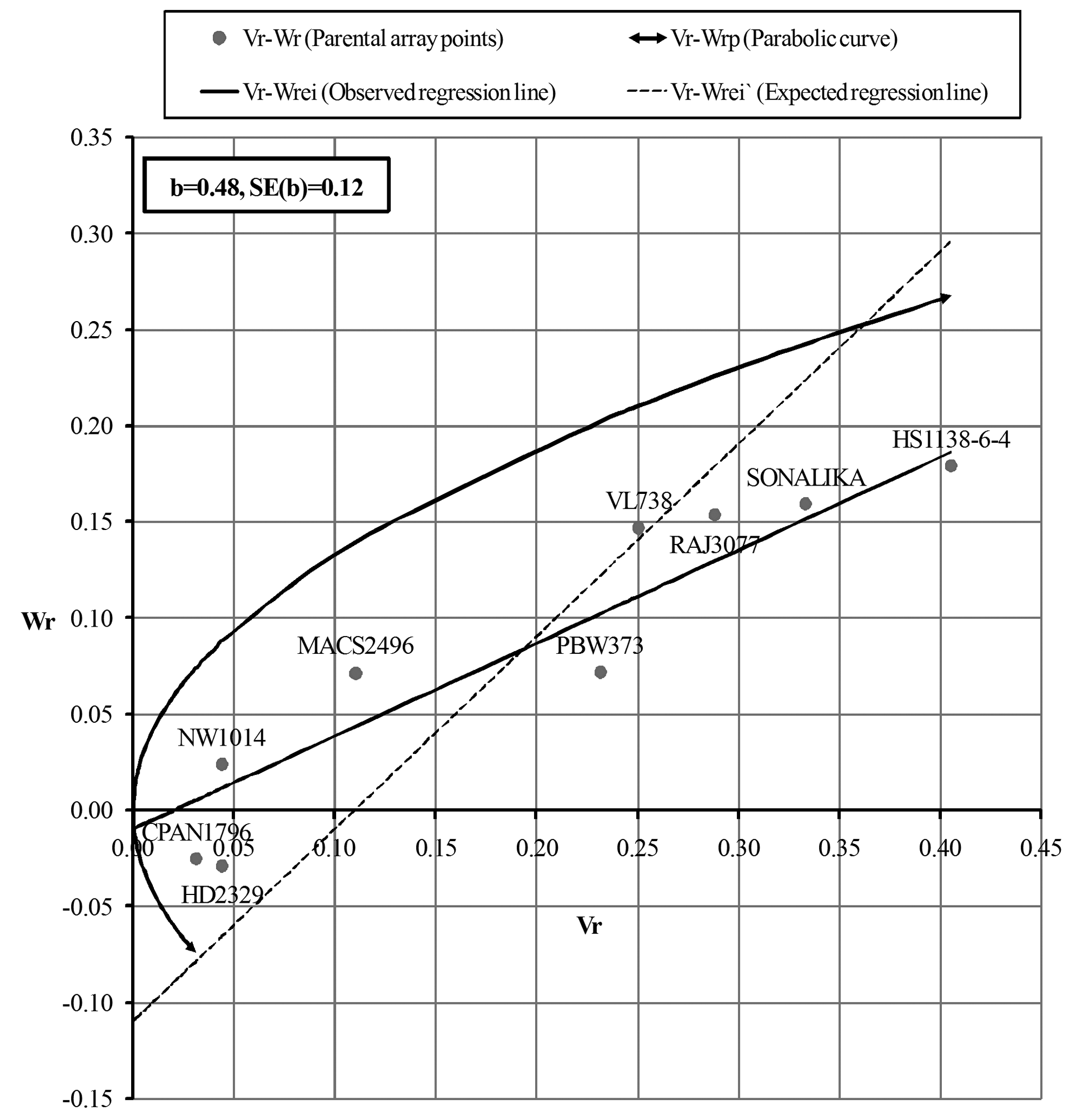
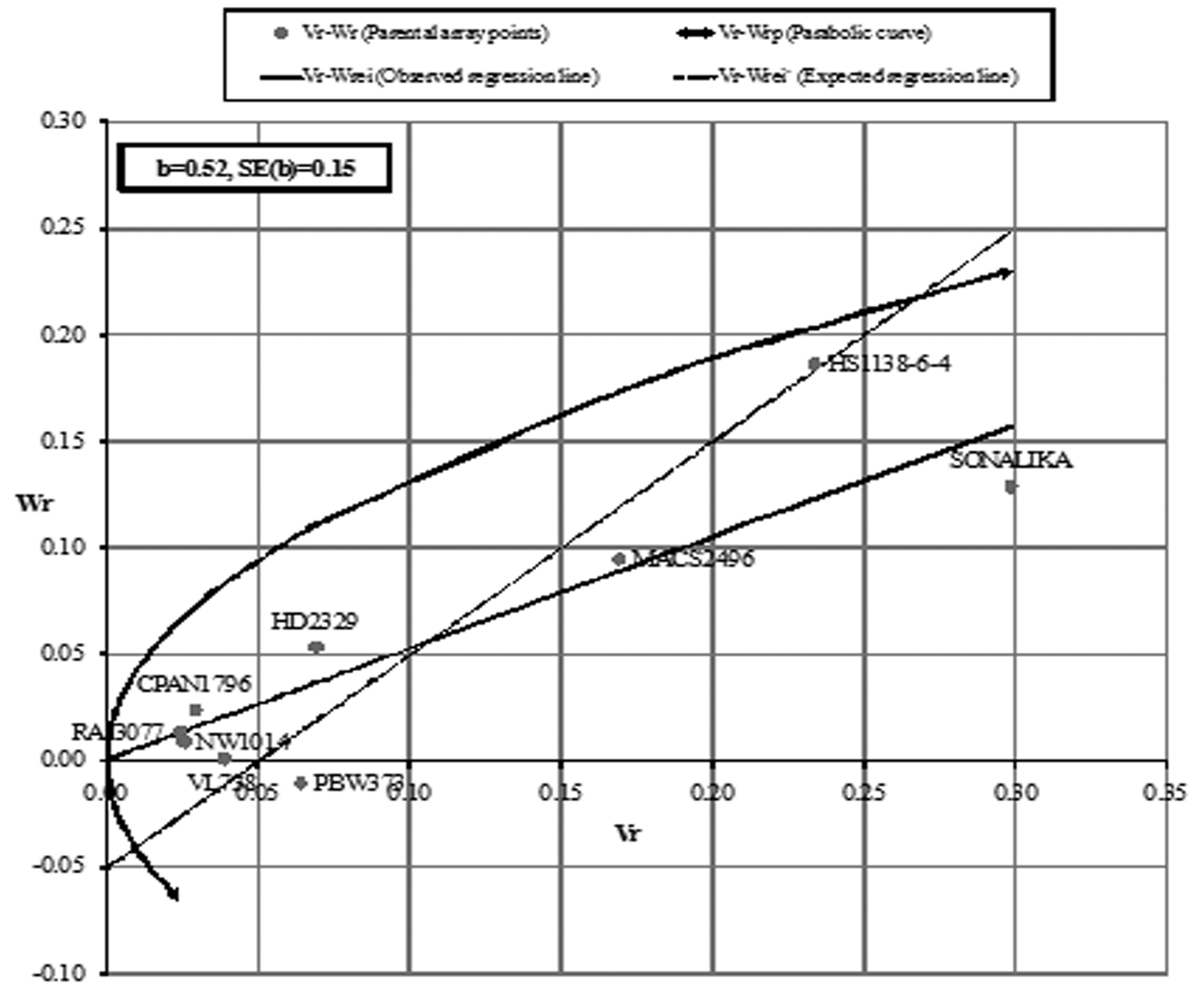
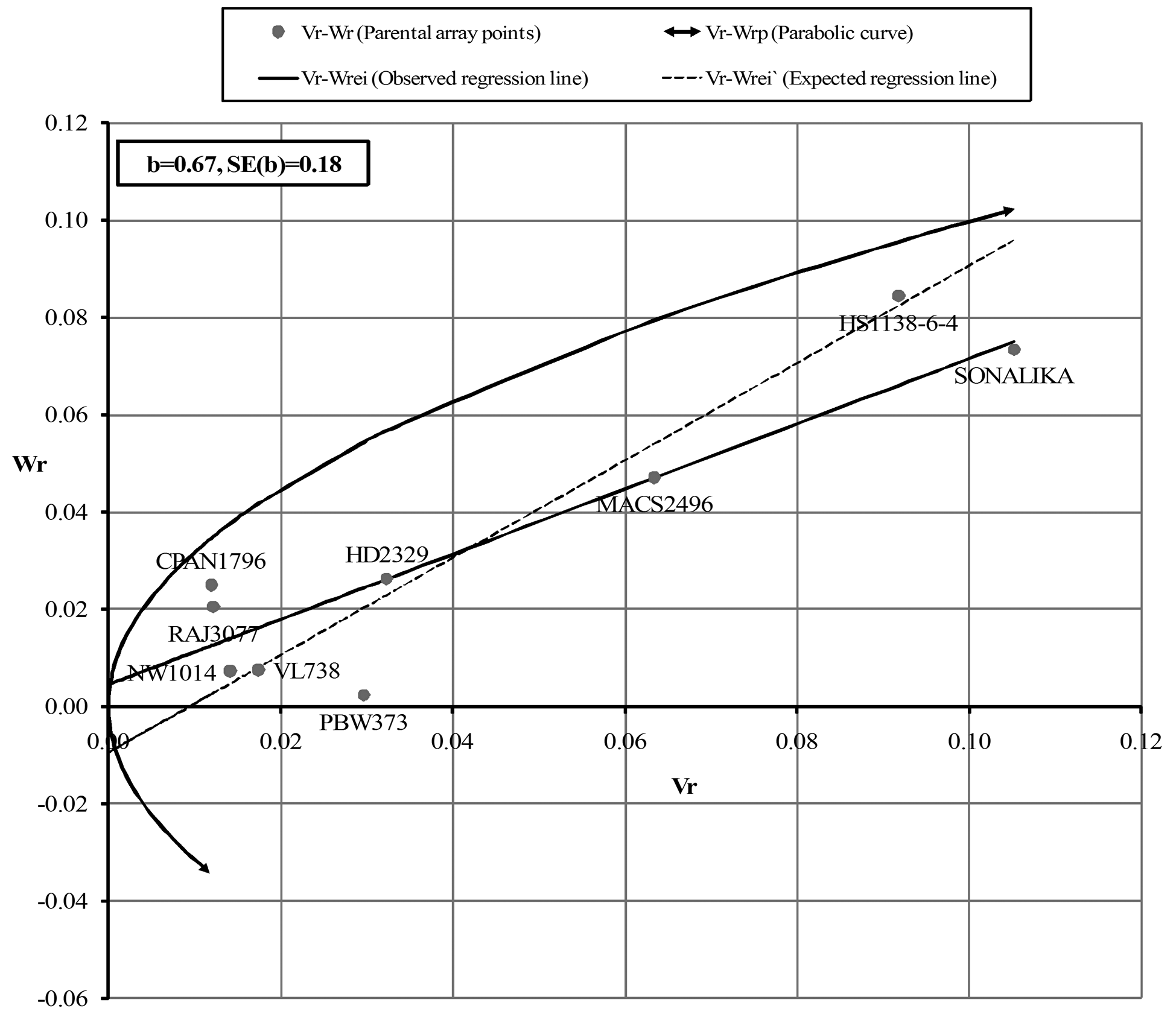
For AUDPC in the F1 (Fig. 1a), the regression line intersected Wr-axis below origin indicating over-dominance, whereas, F2 (Fig. 1b) showed complete dominance. The regression line (Fig. 2a and 2b) intersected Wr-axis just above or close to the point of origin, indicating partial-dominance for FDS in both F1 and F2. Parents CPAN1796, HD2329, NW1014 and MACS2496 exhibited preponderance of minor genes for slow rusting, whereas, SONALIKA and HS1138-6-4 were devoid of such genes for both AUDPC and FDS (F1 and F2).
|
|
.png)
|
|
1.4 Genetic components and its proportions
Analysis of variance for AUDPC and FDS in both F1 and F2 generations are given in Table 2. The difference among treatments was highly significant indicating sufficient variation among them. Components of genetic variation and their proportions are presented in Table 3. Variations due to additive (D), dominance (H1) genetic variance and dominance genetic variance due to + and – effects of genes (H2) as well as net dominance effects h2 as algebraic sum over all loci in heterozygous phase in all cross combinations were significant for AUDPC and FDS (F1 and F2) indicating importance of these components. The average degree of dominance (H1/D)1/2was larger than 1 indicating that each locus was within the range of over-dominance. Proportion of positive and negative genes (H2/4H1)in the parents were 0.19 (AUDPC) and 0.20 (FDS) in F1 and 0.23 in F2 (AUDPC and FDS). The mean covariance of additive and dominance effect of genes F was significant for FDS (F2). The ratio h2/H2 revealed involvement of 1-3 gene(s) in the inheritance of AUDPC and FDS. Preponderance of minor alleles KD/KR was observed in all the cases. Narrow sense heritability (Table 3), was found to be moderate in both F1 and F2 analyses (AUDPC and FDS).
|
|
|
Table 3 Three dimensional scattered diagram based on three PCA scores showing the distribution of different genotypes |
.png)
1.5 Assay of carbohydrate metabolizing enzymes
The analysis of variance of combining ability for AUDPC and FDS showed significant difference in GCA and SCA effects (Table 4). Estimates of GCA and SCA effects have been presented in Table 5. Among the parents, CPAN1796, NW1014, HD2329 and PBW373 were good general combiners, whereas, HS1138-6-4 and SONALIKA were poor general combiners for AUDPC and FDS. However, the susceptible parent HS1138-6-4 had better specific combining ability than SONALIKA with slow rusting lines used. Out of the 36 cross combinations, there were 15 cross combinations that exhibited negative and significant SCA effects for AUDPC and FDS in the F1 and F2 analyses. The cross PBW373 × HS1138-6-4 produced highest magnitude of significant SCA effect. Crosses namely, CPAN1796 × MACS2496, CPAN1796 × HS1138-6-4, CPAN1796 × SONALIKA, NW1014 × MACS2496, NW1014 × SONALIKA, HD2329 × HS1138-6-4, MACS2496 × VL738 and PBW373 × RAJ3077 also had negative and significant SCA effects for both AUDPC and FDS. In general, most of the crosses exhibiting negative significant SCA effects involved good × poor general combiners.
|
|
|
|
.png)
2 Discussions
Five major genes viz, Lr1, Lr10, Lr13, Lr23 and Lr26 were postulated based on seedling reaction test among the parents in different combinations. Slow rusting APR gene Lr34 was detected with the expression of post flowering leaf tip necrosis. Based on pedigree details (Nayar et al., 2001) of different wheat accessions used in the present study we presumed that it was Lr34 in the parents HD2329, PBW373 and VL738 and not other slow rusting genes (Lr46, Lr67 and LrP) which are also linked to Ltn. Major Lr gene effects were nullified by inoculating the experimental material (9 parents, 36 F1s and their corresponding F2s) using a mixture of 10 leaf rust races, that were fully virulent on postulated genes. The race 77-5 virulent on all the postulated genes was also included in the mixture. Low AUDPC and FDS obtained in parents HD2329, PBW373 and VL738 is attributed to the presence of minor genes with additive effects. Since all these parents carry Lr34, remarkable variation in AUDPC (97.10~239.90) and FDS (9.67~20.47) may be due to variation in number of minor genes. More interesting is the fact that parents CPAN1796, NW1014, MACS2496 and RAJ3077 which do not have Lr34 also showed lower AUDPC (85.30~222.60) and FDS (9.57~22.00) as compared to susceptible parents (AUDPC 641.30~723.30; DC 60.67~67.67) strongly suggesting presence of minor genes or modifiers for slow rusting. Apparently, the minor genes in these lines are different to Lr34. Such genes when incorporated with known slow rusting genes like Lr34, Lr46, Lr67 or LrP, can further increase durability of resistance significantly.
The assumptions of diallel analysis was fulfilled in case of FDS (F2) analysis indicating that in rest of the cases the trait was influenced by epistasis and/or non-fulfillment of one or more assumptions. However, the linear relationship between Vr and Wr validates the estimates. The graphical analysis revealed the influence of partial to complete dominance for partial resistance to leaf rust, which is in agreement with Menshawy et al. (2004). The position of parental array points close to regression line revealed the least influence of environment on the inheritance of slow rusting. Parents CPAN1796, HD2329 and NW1014 exhibited preponderance of minor genes with additive effects for both AUDPC and FDS (F1 and F2), whereas, SONALIKA and HS1138-6-4 had absence of such genes attributing to susceptibility.
Contributions of both additive and dominance genetic components were important for the inheritance of AUDPC and FDS (F1 and F2). Estimates of h2 were significantly different from zero in all cases, indicating largely unidirectional dominance in the inheritance of partial resistance to leaf rust. Proportion of positive and negative genes H2/4H1 in the parents were close to 0.25, indicating symmetrical distribution of positive and negative genes for AUDPC and FDS (Menshawy and Youssef, 2004). The ratio h2/H2 revealed involvement of 1-3 genes/groups of minor genes cumulatively exhibiting dominance in the inheritance of AUDPC and FDS. Our estimates of 1 to 3 genes for slow rusting probably includes slow rusting gene Lr34 with large effect and other 2 to 3 genes with minor effects. Das et al. (1993) and Singh et al. (2011) working with slow rusting in wheat have also reported involvement of 4-5 such genes that resulted in ‘near-immunity’ or a high level of resistance along with durability. Studying the inheritance of leaf rust resistance, narrow sense heritability, was found moderate for AUDPC and FDS in both F1 and F2 analyses suggesting the importance of additive gene actions for slow rusting (Das et al., 1992; Kumar et al., 1982; Navabi et al., 2003).
The GCA effects represent fixable genetic components (additive as well as additive × additive gene action), whereas, SCA effects represent non-fixable genetic components. The analysis of combining ability for AUDPC and FDS using F1 and F2 analyses showed genotypes CPAN1796, NW1014, HD2329 and PBW373 were the best general combiners for resistance to leaf rust as they exhibited negative and significant estimates of GCA effects, whereas, parents HS1138-6-4 and SONALIKA, were observed as the poor general combiners for AUDPC and FDS owing to their higher values. These good combiners are expected to produce disease resistant progenies when crossed with other parents. The cross PBW373 × HS1138-6-4 produced highest magnitude of negative significant SCA effect. Crosses namely, CPAN1796 × MACS2496, CPAN1796 × HS1138-6-4, CPAN1796 × SONALIKA, NW1014 × MACS2496, NW1014 × SONALIKA, HD2329 × HS1138-6-4, had desirable SCA effects for MACS2496 × VL738 and PBW373 × RAJ3077 alsoboth AUDPC and FDS. Among the above mentioned crosses exhibiting partial resistance to leaf rust and if their performances were mostly due to non-additive gene effects as revealed by SCA effects would be desirable for slow disease progress only in hybrid generations. Crosses involving good x poor general combiners for wheat leaf rust resistance with desirable negative SCA effects were also reported by Navabi et al. (2003).
On the basis of graphic, component and combining ability analyses of diallel for leaf rust in the present study, it was observed that both AUDPC and FDS were mostly controlled by 1~3 gene(s) having minor effects responsible for slow rusting. Therefore, selection of plants with low to moderate leaf rust severity in early segregating generations would help in identifying plants with minor genes conditioning partial resistance to leaf rust. Moderate narrow sense heritability estimates suggest that success in selecting and fixing desirable lines with partial resistance to leaf rust in early segregating generations can be easily achieved.
3 Materials and methods
3.1 Gene postulation in parental materials
The seedling reaction test of the parents was done using specific leaf rust races for postulating the major genes in the parents at Directorate of Wheat Research, (DWR) Regional Station, Flowerdale, Shimla (India) by spraying 75 mg of urediniospores in 250 ml light weight mineral oil ‘Soltrol 170’ on 7~8 days old seedlings with the help of an atomizer. The inoculated material was kept for 1 hour to allow the oil to evaporate. Later, the inoculated plants were sprayed with a fine mist of water and kept for 48 hours in water saturated chamber at 18±20℃ for incubation facilitating spore germination and penetration. The inoculated material was then transferred to the glasshouse benches (maintained at 23±2℃). The infection types produced after 13~14 days were recorded according to Nayar et al. (1994). The principle used for gene postulation was based on gene-for-gene specificity (Flor, 1971). The infection types produced by a diverse group of P. triticina isolates on the parental lines were compared with infection types produced by the same isolates on standard differential sets of near-isogenic Thatcher wheat lines.
The infection types were categorized as per Nayar et al. (1994). For example, naught fleck (0;) = immune, no visible infection; fleck minus (;-) = nearly immune, micro flecking visible; fleck (;) = very resistance, no uredia but hypersensitive fleck; one (1) = very resistance, uredia minute, surrounded by distinct necrotic areas; two (2) = moderately resistant, uredia small to medium, surrounded by chlorotic or necrotic border; three (3) = moderately susceptible, uredia small to medium in size, chlorotic areas may be present; three three plus (33+) = susceptible, no chorotic or necrosis, uredia profusely sporulating; three plus/ four (3+/4) = highly susceptible, no chlorosis or necrosis, uredia profusely sporulating, ring may be formed. Designation of + and – were added to 0 to 4 infection type to indicate large and smaller uredia than normal, respectively. Generally, infection types 0 to 2+ were considered as low infection type and 3 to 4 were considered as high infection type.
3.2 Diallel mating among parental materials
The parental material consisted of slow rusting (CPAN1796, NW1014, HD2329, MACS2496, PBW373, RAJ3077 and VL738) and susceptible parents (HS1138-6-4 and SONALIKA). Crosses were made in diallel fashion excluding reciprocals to obtain 36 F1s by hand-emasculation and artificial pollination using “go-go” method (Thomas and Anderson, 1978). Thirty six F1s were grown to obtain F2 seeds at Indian Agricultural Research Institute (IARI) Regional Station, Wellington, Tamil Nadu (India).
3.3 Adult Plant Reaction (APR) test using mixture of virulent leaf rust races
Nine parental lines, 36 F1s and their F2s were evaluated in randomized complete block design with three replications maintaining row to row and plant to plant spacing of 25 and 10 cm, respectively at Agricultural Research Farm, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi (India) during the Winter 2008-09 crop season. One row of the highly susceptible variety ‘Agra Local’ as spreader on both sides of the each plot and three rows around the experiment were planted. Artificial inoculation was done with the mixture of virulent leaf rust races viz, 12-2, 77-2, 77-5, 104-1, 104-2, 104-3, 107-1, 108, 108-1 and 162 (to nullify the effect of major genes present in parents/ F1s/ F2s and determine the effect of minor genes) obtained from Flowerdale, Shimla by spraying the spreader rows as well as nine parental lines, 36 F1s and their F2s with urediniospores suspended in a light-weight mineral oil at growth stage 35 (GS 35) as per Zadoks et al. (1974).
3.4 Assessment of leaf rust severity
The parents, F1s and F2s plants were assessed for average leaf rust severity based on the infection at the flag and penultimate leaves of the two most advanced tillers of each plant. Severities were scored on 10 plants in case of parents and F1s and 75 plants in each F2s in each replication. The first scoring of severity was done when the leaf rust severity of the spreader cultivar ‘Agra Local’ reached 80 to 100% at GS 50 and continued up to GS 87 of test materials. Observations on rust severities were recorded visually according to modified Cobb’s scale (Peterson et al., 1948) 5 times at an interval of 7 days and area under disease progress curve (AUDPC) was calculated following Das et al. (1992). The rust severity values of the fifth reading were considered as final disease severity (FDS). The AUDPC and FDS values were transformed by log10 transformation to normalize the data.
3.5 Genetic analysis
Analysis of variance was carried out for AUDPC and FDS, separately for the F1 and F2 following Panse and Sukhatme (1967). The array variance (Vr) and parent-offspring covariance (Wr) were computed for graphical and component analyses according to Hayman (Hayman, 1954) in F1 and Jinks (Jinks, 1956) in F2. The Wr values were regressed on Vr values and relationship was plotted to make the Vr-Wr graph. Combining ability analysis was carried out following the Model I and Method II of Griffing (1956). Narrow-sense heritability was estimated separately for each generation as the ratio of additive and/or additive × additive genetic variance to phenotypic variance. In the F2, it was calculated following Verhalen and Murray (1969).
Acknowledgements
The first author gratefully acknowledges financial assistance provided by the Banaras Hindu University (BHU), infrastructure facilities by the Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi (India); DWR Regional Station, Flowerdale, Shimla (India) and IARI Regional Station, Wellington, Tamil Nadu (India) for completion of the research work. The first author thankfully acknowledges the help received during manuscript preparation from Dr AK Singh, Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi (India).
Reference
Caldwell R.M., 1968, Breeding for general and/ or specific plant disease resistance. In: Shepherd KW (ed) International wheat genetic symposium, 3rd ed. Academy of Science, Canberra
Das M.K., Rajaram S., Mundt C.C., and Kronstad W.E., 1992, Inheritance of slow rusting to leaf rust in wheat, Crop Sci., 32:1452-1456
http://dx.doi.org/10.2135/cropsci1992.0011183X003200060028x
Das M.K., Rajaram S., Kronstad W.E., Mundt C.C., and Singh R.P., 1993, Associations and genetics of three components of slow rusting in leaf rust of wheat, Euphytica, 68: 99-109
http://dx.doi.org/10.1007/BF00024159
Flor H.H., 1971, Current status of the gene-for-gene concept, Annu. Rev. Phytopathol., 9:275-296
http://dx.doi.org/10.1146/annurev.py.09.090171.001423
Griffing B., 1956, Concept of general and specific combining ability in relation to diallel crossing systems, Aust. J. Biol. Sci., 9: 463-493
Hayman B.I., 1954, The theory and analysis of diallel crosses, Genetics, 43: 789-809
Herrera-Foessel S.A., Lagudah E.S., Huerta-Espino J., Hayden M.J., Bariana H.S., Singh D., and Singh R.P., 2011, New slow rusting leaf rust and stripe rust resistance genes Lr67 and Yr46 in wheat are pleiotropic or closely linked, Theor. Appl. Genet., 122: 239-249
http://dx.doi.org/10.1007/s00122-010-1439-x
Hiebert C.W., Thomas J.B., McCallum B.D., Humphreys D.G., DePauw R.M., Hayden M.J., Mago R., Schnippenkoetter W., and Spielmeyer W., 2010, An introgression of wheat chromosome 4DL in RL6077 (Thatcher*6/PI 250413) confers adult plant resistance to stripe rust and leaf rust (Lr67), Theor. Appl. Genet., 121: 1083-1091
http://dx.doi.org/10.1007/s00122-010-1373-y
Jinks J.L., 1956, The F2 and backcross generations from a set of diallel crosses, Heredity, 10: 1-30
http://dx.doi.org/10.1038/hdy.1956.1
Knott D.R., 1982, Multigenic inheritance of stem rust resistance in wheat, Crop. Sci., 22(2): 393-399
http://dx.doi.org/10.2135/cropsci1982.0011183X002200020045x
Krattinger S.G., Lagudah E.S., Spielmeyer W., Singh R.P., Huerta-Espino J., McFadden H., Bossolini E., Selter L.L., and Keller B., 2009, A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat, Science, 323: 1360-1363
http://dx.doi.org/10.1126/science.1166453
Kuhn R.C., Ohm H.W., and Shaner G., 1980, Inheritance of slow leaf-rusting resistance in Suwon 85 wheat, Crop. Sci., 20: 655-659
http://dx.doi.org/10.2135/cropsci1980.0011183X002000050028x
Kumar S., Singh R.M., and Singh R.B., 1982, Inheritance of slow development of brown rust in wheat, Indian J. Agric. Sci., 52(5): 324-330
Menshawy A.M.M., and Youssef W.A., 2004, Inheritance of resistance to stripe and leaf rusts in spring wheat, Egypt J. Agri. Res., 82(3): 1199-1211
Navabi A., Singh R.P., Tewari J.P., and Briggs K.G., 2003, Genetic analysis of adult-plant resistance to leaf rust in five spring wheat genotypes, Plant Dis., 87: 1522-1529
http://dx.doi.org/10.1094/PDIS.2003.87.12.1522
Nayar S.K., Nagarajan S., Prashar M., Bhardwaj S.C., Jain S.K., and Datta D., 2001, Revised catalogue of genes that accord resistance to Puccinia species in wheat, Research Bulletin No. 3: 48 pp. Directorate of Wheat Research, Regional Station, Flowerdale, Shimla 171002 (India)
Nayar S.K., Tandon J.P., Kumar J., Prashar M., Bhardwaj S.C., Goel L.B., and Nagarajan S., 1994, Basis of rust resistance in Indian wheat, Research Bullelin No. 1: 32 pp. Directorate of Wheat Research, Regional Station, Flowerdale, Shimla 171002 (India)
Panse V.G., and Sukhatme P.V., 1967, Statistical methods for agricultural workers, 2nd edn. Indian Council of Agricultural Research, New Delhi
Peterson R.F., Campbell A.B., and Hannah A.E., 1948, A diagrammatic scale for estimating rust severity on leaves and stems of cereals, Can. J. Res. C., 26: 496-500
http://dx.doi.org/10.1139/cjr48c-033
Roelfs A.P., Singh R.P., and Saari E.E., 1992, Rust Diseases of Wheat: Concepts and Methods of Disease Management. CIMMYT, Mexico, DF CIMMYT, pp.81
Samborski D.J., 1984, Wheat leaf rust. In: Roelfs AP (ed) The Cereal Rusts, Vol. 2. WR Bushnell, Academic Press Inc, Orlando, FL, pp.39-60
Shaner G., Buechley G., and Nyquist W.E., 1997, Inheritance of latent period of Puccinia recondita in wheat, Crop Sci., 37: 748-756
http://dx.doi.org/10.2135/cropsci1997.0011183X003700030010x
Singh R.P., Huerta-Espino J., Bhavani S., Herrera-Foessel S.A., Singh D., Singh P.K., Velu G., Mason R.E., Jin Y., Njau P., and Crossa J., 2011, Race non-specific resistance to rust disease in CIMMYT spring wheats, Euphytica, 179: 175-186
http://dx.doi.org/10.1007/s10681-010-0322-9
Singh R.P., Huerta-Espino J., and William M., 2001, Slow rusting genes based resistance to leaf and yellow rusts in wheat: genetics and breeding at CIMMYT, pp.103-108 In: Proc. 10th Assem. Wheat Breed. Soc. Australia, Mildura, Australia. Wheat Breed. Soc. Aust. Inc., Mildura, Australia
Smale M., Singh R.P., Sayre K., Pingali P., Rajaram S., and Dubin H.J., 1998, Estimating the economic impact of breeding nonspecific resistance to leaf rust in modern bread wheats, Plant Dis., 82:1055-1061
http://dx.doi.org/10.1094/PDIS.1998.82.9.1055
Thomas J.B., and Anderson R.G., 1978, Cross incompatibility of common wheat with rye: effect of varying the day of pollination on seed set and development, Can. J. Bot., 56: 3000-3007
http://dx.doi.org/10.1139/b78-364
Verhalen L.M., and Murray J.C., 1969, A diallel analysis of several fibre property traits in upland cotton (Gossypium hirsutum L.) II, Crop Sci., 9(3):311-315
http://dx.doi.org/10.2135/cropsci1969.0011183X000900030018x
William M., Singh R.P., Huerta-Espino J., Ortiz Islas S., and Hoisington D., 2003, Molecular marker mapping of leaf rust resistance gene Lr46 and its association with stripe rust resistance gene Yr29 in wheat, Phytopathol., 93: 153-159
http://dx.doi.org/10.1094/PHYTO.2003.93.2.153
Zadoks J.C., Chang T.T., and Konzak C.F., 1974, A decimal code for the growth stages of cereals, Weed Res., 14:415-421
http://dx.doi.org/10.1111/j.1365-3180.1974.tb01084.x



. PDF(252KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Manoj Kumar
. Rishi Singh
. Mohinder Prashar
. Rajiva Agrawal
. Ramesh Chand
Related articles
. AUDPC
. Diallel
. FDS
. Leaf rust
. Partial resistance
. Wheat
Tools
. Email to a friend
. Post a comment