Research Article
Exogenous Glutathione Improves Salinity Stress Tolerance in Rice (Oryza sativa L.) 
2 Department of Botany, University of Otago, New Zealand
 Author
Author  Correspondence author
Correspondence author
Plant Gene and Trait, 2016, Vol. 7, No. 11 doi: 10.5376/pgt.2016.07.0011
Received: 09 Aug., 2016 Accepted: 29 Sep., 2016 Published: 03 Nov., 2016
Hussain B.M.N., Akram S., Raffi S.A., Burritt D.J., and Hossain M.A., 2016, Exogenous Glutathione Improves Salinity Stress Tolerance in Rice (Oryza sativa L.), Plant Gene and Trait, 7(11): 1-17 (doi: 10.5376/pgt.2016.07.0011)
The effect of salt stress and the beneficial roles of exogenous glutathione (GSH) in modulating salt stress tolerance in rice (Oryza sativa L.) were investigated by conducting an experiment in the net house of the Department of Genetics and Plant Breeding, Bangladesh Agricultural University, Mymensingh. The experiment used 11 diverse rice genotypes and used a Randomized Complete Block Design with four replicates and three treatments, a control, salt (200 mM NaCl) and salt + 2mM GSH. Analysis of variance for yield and yield contributing traits showed significant (p< 0.01) variation among the genotypes. Significant G (genotype) × T (treatment) interaction was found for the following traits, days to maturity, plant height, number of unfilled grains per panicle and spikelet fertility. Exposure to salt stress at reproductive stage resulted in significant decreases in yield and yield attributing traits, as compared to the control, with greater reductions observed in salt sensitive and salt non-tolerant genotypes. Exogenous GSH application to salt stressed plants resulted in increased yield and yield attributing traits, and the number of unfilled grains per panicle was reduced in most genotypes, compared to plants subjected to salt stress without GSH. The positive influence of exogenous GSH application was most pronounced for salt-stressed plants of salt sensitive and non-tolerant genotypes as compared to the tolerant genotypes or advanced breeding lines. The present study showed that the application of GSH can improve salt stress tolerance of salt sensitive rice plants and positively influence yield contributing traits.
Background
Salt stress is one of the most severe constraints to crop productivity worldwide, and thus salinization of water and soils in major agricultural areas has become a major concern for global food security. It is estimated that more than 6% of the world’s land and 30% of irrigated areas already suffer from salinity problems (Chaves et al., 2009). The negative impact of salinity on crop cultivation has been well documented in the coastal areas of Bangladesh and predicated global climate change will intensify the frequency and duration of such abiotic stresses (Karim et al., 1990; Uddin, 2005; Dasgupta et al., 2014). Although a few salt tolerant rice varieties have been developed, by the Bangladesh Rice Research Institute (BRRI) and Bangladesh Institute of Nuclear Agriculture (BINA), their yields under saline conditions are comparatively low, compared to the yields of varieties grown under non-saline conditions. Hence, there is an urgent need to develop rice varieties that can withstand high levels of salt stress. To facilitate the development of cultivars that can survive and produce high yields under saline conditions, a thorough understanding of the various morphological, physiological and molecular mechanisms that govern yield under salt stress conditions is required.
Rice (Oryza sativa L.) is one of the most important crop plants, providing food for more than half of the world’s population (Malik et al., 2008). Rice yields decrease 12% for every unit (dSm-1) increase in electrical conductivity (EC) above 3 dSm-1 (Mass and Grattan, 1999). Reductions in grain yields for rice is much greater when plants are subjected to salt stress at the reproductive stage, as compared to vegetative stage (Akbar et al., 1972; Zeng et al., 2000; Hossain et al., 2015) and the reduction in yield depends on the salinity level and the duration of exposure to high salinity. Salinity has the greatest negative impacts on panicle initiation, spikelet formation, pollen germination and fertilization, with significant effects was also observed on panicle weight, panicle length, the number of primary branches per panicle, the number of unfilled grains per panicle, the total number of seeds per panicle, total seed weight per panicle, 1000-seed weight and total seeds per plant (Abdullah et al., 2002; Lauchli and Grattan, 2007). Exposure to excess salt causes both ionic and osmotic stress in plants (Kader and Lindberg, 2010). In addition to ionic and osmotic stress, one of the most important biochemical changes observed in plants under salt stress is the abrupt increased in reactive oxygen species (ROS), such as hydrogen peroxide, superoxide and hydroxyl radical that ultimately leads to oxidative damage (Hossain et al., 2009, 2013a, 2013b; Upadhaya et al., 2011; Yamme et al., 2012; Ghosh et al., 2014; Nahar et al., 2015a; Mostofa et al., 2015a). Methylglyoxal (MG), which can be detoxified by the glyoxalase system, is also overproduced in plants under salt stress and this can also result in increased oxidative damage (Hossain et al., 2009; Hoque et al., 2010). Pollen and spikelet fertility and pollen germination are the most important determinates of rice yields under salt stress (Zeng et al., 2000; Abdullah et al., 2002; Hasanuzzaman et al., 2009). Drought induced spikelet sterility was reported to be associated with an abrupt increase in ROS and/or inefficient antioxidant defense systems in rice (Selote and Khanna-Chopra, 2004). Therefore, breeding for salt stress requires the development of rice varieties with high pollen and spikelet fertility, and higher ROS and MG detoxification capacities. Enhanced ROS and MG detoxification capacities are among the most important determinants of salt stress tolerance and changes in the activities of antioxidant enzymes in response to salinity have been reported to differ between salt tolerant and sensitive cultivars of various crop plants (Sairam et al., 2002; Meloni et al., 2003; El-Shabrawi et al., 2010).
Glutathione (GSH) is one of the most abundant water soluble antioxidant molecules and plays important stress defensive roles in plants, including ROS detoxification and cellular signalling that triggers adaptive responses in plants under various abiotic stresses including salt stress (Labudda et al., 2014; Nahar et al., 2015a, 2015b; Mostofa et al., 2015a). Reduced GSH can directly detoxify ROS and regulate the activities of GSH-dependent ROS and MG detoxifying enzymes (Hossain et al., 2013a; Ramírez et al., 2013; Nahar et al., 2015a, b; Mostofa et al., 2014, 2015a,b). It is a pivotal component of the ascorbate-glutathione (AsA-GSH) cycle, a system that controls H2O2 levels in plant cells. Plants usually maintain a high cellular GSH/GSSG ratio, but under stressful conditions GSH reacts with oxidants and is converted to GSSG, leading to a lower GSH/GSSG ratio that can result in oxidative damage (Hossain et al., 2013c; Nahar et al., 2015a, b; Mostofa et al., 2015a). Under stressful conditions GSH levels decrease in stress susceptible genotypes, but in contrast increase in stress tolerant genotypes and this concomitant increase in GSH levels has been correlated to drought or heat-induced oxidative stress tolerance (Selote and Khanna-Chopra, 2004; Nahar et al., 2015a). Additionally, the beneficial roles of exogenous GSH in improving nutrient deficiency and heavy metal stress tolerance in various plant species have been reported in several studies, where GSH was found to influence ROS metabolism as well as nutrient or metal uptake and utilization (Cai et al., 2011a, 2011b; Chen et al., 2012; Ramírez et al., 2013; Mostofa et al., 2014). Recently, it has been reported that exogenous GSH improved salt tolerance by increasing ROS and MG detoxification, by modulating antioxidant enzyme activities and the activities of glyoxalase enzymes (Wang et al., 2014; Nahar et al., 2015a). In addition, transgenic rice plants over-expressing GSH biosynthetic genes had higher salinity and methyl viologen (MV) induced oxidative stress tolerance than non-transgenic control plants (Choe et al., 2013). Therefore, the assessment of popular salt tolerant and high yielding rice genotypes, tolerant advanced breeding lines and sensitive genotypes under salt stress, as well as the possible mechanisms of GSH-induced salt stress tolerance, are needed to understand in-depth mechanisms involved in salt tolerance and to breed rice varieties with higher salt tolerance. Importantly, most previous studies using exogenous GSH applications were conducted under hydroponic conditions at various stages of seedling growth. To the best of our knowledge, there are no reports on the beneficial roles of exogenous GSH application as a means to improve salinity stress tolerance at the reproductive stage. The present research was conducted to evaluate the effects of salt stress and explore the protective roles of exogenous GSH in modulating salt tolerance at the reproductive stage in rice, by analysing yield and yield-attributing traits.
1 Results
1.1 Analysis of variance
A two-way analysis of variance for all the characters (viz., days to maturity, number of effective tillers per plant, plant height, panicle length, number of filled grains per panicle, number of unfilled grains per panicle, spikelet fertility, 100-seed weight, yield per panicle) showed highly significant (p< 0.01) variation among the genotypes studied. In case of treatment, all the characters except days to maturity showed significant (p< 0.01) variation. For the G × T (genotype × treatment) interaction, days to maturity, plant height, number of unfilled grains per panicle, spikelet fertility were found to be significant (p< 0.01) (Table 1). The mean sum of squares of all the characters are presented in Table 1.
1.2 Effect of salt stress and salt+2mM GSH treatments on rice varieties
1.2.1 Days to maturity
The mean number of days required to achieve maturity among all the genotypes was 123.20 days (Table 2a). With a range of 104.8 days (BRRI dhan56) to 149 days (BRRI dhan41) under control conditions, 105.3 days (BRRI dhan56) to 149.5 days (BRRI dhan41) under salt stress and 105.0 (BRRI dhan56) days to 149.5 days (BRRI dhan41) under salt stress + 2mM GSH (Table 2). Salt stress and exogenous GSH application had no significant effect on days to maturity for most of the genotypes tested. However, a G × T interaction for this trait showed significant (p< 0.01) variation (Table 3).
1.2.2 Number of effective tillers per plant
The highest number (18.63) of effective tillers was found in Binadhan-7, under control conditions and the lowest (9.75) was found in FL-478 under salt stress (Table 2a). Imposition of salt stress resulted in a significant decrease in the number of effective tillers per plant, with the greatest reduction (23.17%) recorded for genotype Binadhan-7 followed by FL-478 (21.79%), Binadhan-8 (18.61%), BRRI dhan56 (17.85%), Nona Bokra (16.13%), BRRI dhan53 (7.06%), IR29 (5.76%), BRRI dhan41 (5.44%), BRRI dhan47 (3.61%), Binadhan-10 (2.91%), and the least reduction (2.04%) was observed in BRRI dhan40 (Table 3; Figure 1), as compared to control. Exogenous GSH resulted in a significant increase in the number of effective tillers per plant, with the greatest increase (16.33%) found in BRRI dhan56 followed by FL-478 (14.29%), BRRI dhan53 (11.46%), Binadhan-7 (11.03%), Binadhan-8 (10.42%), BRRI dhan41 (8.00%), IR29 (5.44%), BRRI dhan40 (2.97%) and the least increase (2.35%) in BRRI dhan47 (Figure 1), as compared to plants subjected to salt stress only. However, the number of effective tillers per plant for Binadhan-10 & Nona Bokra was not greatly affected by GSH application (Table 3; Figure 1).
.png) Table 1 Analysis of variance of morphological characters of 11 rice genotypes grown under control, salt and salt + 2mM GSH conditions |
 Table 2a Mean performances of 11 rice genotypes based on different morphological traits related to yield grown under control, salt and salt + 2mM GSH conditions |
.png) Figure 1 The percent change of the number of effective tillers per plant in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.3 Plant height (cm)
The highest (123.2 cm) plant height was for genotype Nona Bokra under control conditions and the lowest (80.56 cm) for genotype Binadhan-7 under salt +2mM GSH (Table 2a). The G × T interaction for this trait showed significant (p< 0.01) variation (Table 3). Salt stress led to a significant reduction in plant height, as compared to controls, the greatest reduction (7.67%) was for genotype BRRI dhan56, followed by BRRI dhan40 (7.18%), BRRI dhan41 (6.52%), BRRI dhan53 (5.55%), IR29 (5.06%), Nona Bokra (4.17%), FL-478 (3.99%) and Binadhan-8 (1.50%), with little reduction in growth (0.91%) was observed for Binadhan-7. Binadhan-10 was not affected by salt application (Figure 2). Application of exogenous GSH resulted in increased plant heights compared to plants under salt stress, the greatest increase in plant height (7.29%) was found for BRRI dhan56 followed by BRRI dhan41 (6.85%), FL-478 (4.99%), BRRI dhan40 (4.79%), Nona Bokra (3.64%), IR29 (2.46%), Binadhan-10 (1.93%) and the least reduction (1.57%) in plant height was for BRRI dhan47 (Figure 2). Moreover, the application of GSH did not significantly influence plant height for the genotypes of Binadhan-7, Binadhan-8 and BRRI dhan53.
.png) Figure 2 The percent change of plant height (cm) in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.4 Panicle length (cm)
The highest panicle length (26.01 cm) was for Binadhan-10 under salt + 2 mM GSH and the lowest (20.05 cm) for Nona Bokra under salt stress (Table 2a). The G × T interaction was not significant for this trait (Table 3). Imposition of salt stress reduced panicle length, the greatest reduction (14.66%) was for the genotype Nona Bokra followed by BRRI dhan40 (10.79%), Binadhan-7 (7.12%), FL-478 (4.31%), BRRI dhan41 (4.18%), BRRI dhan56 (3.59%), Binadhan-8 (2.83%), IR29 (1.92%), BRRI dhan53 (1.31%), Binadhan-10 (1.00%), and the least reduction (0.91%) was observed for BRRI dhan47 (Figure 3). Exogenous GSH caused a significant increase, panicle length, with the greatest increase (10.24%) recorded for Nona Bokra followed by Binadhan-7 (6.98%), BRRI dhan41 (6.72%), BRRI dhan40 (5.98%), BRRI dhan56 (5.07%), Binadhan-8 (4.48%), FL-478 (2.25%), BRRI dhan47 (1.71%), and with only a 1.14% increase for Binadhan-10 (Figure 3), compared to the plants treated with salt only. However, GSH had no significant effect on panicle length for the genotypes BRRI dhan53 and IR29 (Table 3; Figure 3).
.png) Figure 3 The percent change of panicle length (cm) in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.5 Number of filled grains per panicle
The highest number (120.60) of filled grains per panicle was found for Nona Bokra under control conditions and the lowest number (70.78) of filled grains per panicle was found for IR29 under salt stress (Table 2b). A significant decrease in the number of filled grains per panicle in response to salt stress was observed, with the greatest reduction (42.14%) for the genotype IR29, followed by Nona Bokra (28.58%), BRRI dhan40 (27.0%), BRRI dhan53 (24.79%), BRRI dhan41 (23.39%), FL-478 (21.68%), BRRI dhan56 (21.45%), Binadhan-10 (21.41%), Binadhan-7 (17.26%), BRRI dhan47 (11.51%), and Binadhan-8 (10.90%) (Figure 4). For salt + 2mM GSH, a significant increase in the number of filled grains per panicle was observed, the greatest increased (20.45%) was for Nona Bokra, followed by BRRI dhan40 (20.34%), IR29 (18.44%), BRRI dhan53 (17.10%), BRRI dhan56 (14.91%), BRRIdhan41 (13.35%), Binadhan-10 (11.27%), Binadhan-7 (10.35%), FL-478 (9.58%), Binadhan-8 (8.87%), and 3.66% for BRRI dhan47 (Table 3; Figure 4).
 Table 2b Mean performances of 11 rice genotypes based on different morphological traits related to yield grown under control, salt and salt + 2mM GSH conditions (cont’d) |
.png) Table 3 Combined effect of genotypes and treatments interaction on yield and yield contributing traits of rice |
.png) Figure 4 The percent change of number of filled grains per panicle in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.6 Number of unfilled grains per panicle
BRRI dhan56 showed the highest number (57.72) of unfilled grains under salt conditions, with the lowest (21.43%) found for genotype BRRI dhan53 under salt + 2mM GSH conditions (Table 2b). The G × T interaction for this trait showed significant (p< 0.01) variation (Table 3). Imposition of salt stress resulted in a significant increase in the number of unfilled grains per panicle, with 43.21% recorded for BRRI dhan56 followed by IR29 (43.06%), Binadhan-10 (37.49%), Nona Bokra (30.46%), Binadhan-7 (30.31%), BRRI dhan47 (25.14%), Binadhan-8 (24.76%), BRRI dhan40 (23.89%), BRRI dhan53 (22.86%), FL-478 (7.29%) and 1.62% for BRRI dhan41 (Figure 5), as compared to control condition. GSH had significant reduction in the number of unfilled grains per panicle, with a 56.53% reduction for IR29 followed by Binadhan-7 (36.37%), BRRI dhan53 (35.69%), Nona Bokra (35.16%), BRRI dhan56 (34.43%), Binadhan-10 (25.69%), BRRI dhan47 (21.79%), Binadhan-8 (21.59%), BRRI dhan40 (13.92%) and 1.88% for FL-478, as compared to salt stress. The salt + 2mM GSH treatment did not influence genotype BRRI dhan41 with respect to this character (Figure 5).
.png) Figure 5 The percent change of number of unfilled grains per panicle in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.7 Spikelet fertility
Binadhan-10 showed the highest (83.41%) spikelet fertility under control conditions whereas the lowest spikelet fertility (62.34%) under salt stress condition was found for BRRI dhan56 (Table 2b). The G × T interaction regarding this trait showed significant (p< 0.01) variation (Table 3). Imposition of salt stress resulted in a significant decrease in spikelet fertility, the greatest reduction (28.68%) was recorded for genotype IR29 followed by BRRI dhan56 (25.00%), Binadhan-10 (15.98%), Nona Bokra (14.48%), Binadhan-7 (13.83%), BRRI dhan40 (11.44%), BRRI dhan53 (10.94%), BRRI dhan47 (10.19%), Binadhan-8 (9.02%), FL-478 (6.05%) and (5.30%) for BRRI dhan41 (Figure 6), as compared to controls. GSH had significant effect, increasing spikelet fertility under salt stress conditions, with the greatest increase (17.74%) recorded for IR29 followed by BRRI dhan56 (13.86%), Nona Bokra (11.30%), Binadhan-7 (10.29%), BRRI dhan53 (10.06%), Binadhan-10 (8.39%), BRRI dhan40 (8.00%), Binadhan-8 (6.47%), BRRI dhan47 (5.86%), BRRI dhan41 (2.78%), with 2.74% for FL-478 (Table 3; Figure 6) as compared to the plants treated with salt only.
.png) Figure 6 The percent change of spikelet fertility in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.8 100-seed weight (g)
The highest 100-seed weight (2.90g) was found for Binadhan-10 under control conditions and the lowest 100-seed weight (2.22g) was found for IR29 under salt stress conditions (Table 2b). A significant reduction in 100-seed weight was observed in response to salt stress, the greatest reduction (8.20%) was recorded for BRRI dhan41 followed by Nona Bokra (7.47%), Binadhan-10 (7.14%), BRRI dhan40 (6.56%), BRRI dhan47 (6.12%), BRRI dhan53 (5.52%), IR29 (5.42%), BRRI dhan56 (5.11%), Binadhan-8 (4.06%), Binadhan-7 (2.90%), with 1.78% for FL-478 (Table 3; Figure 7), as compared to controls. Treatment 2mM GSH under salt stressed conditions increased 100-seed weight, the greatest increase in 100-seed weight (6.08%) was for BRRI dhan41, followed by BRRI dhan40 (5.54%), IR29 (3.91%), Nona Bokra (3.79%), FL-478 (1.51%), BRRI dhan56 (1.47%), Binadhan-7 (0.54%), Binadhan-10 (0.48%), with 0.041% for BRRI dhan47. GSH treatment did not greatly influence the 100-seed weight of the genotypes Binadhan-8 and BRRI dhan53 (Table 3; Figure 7).
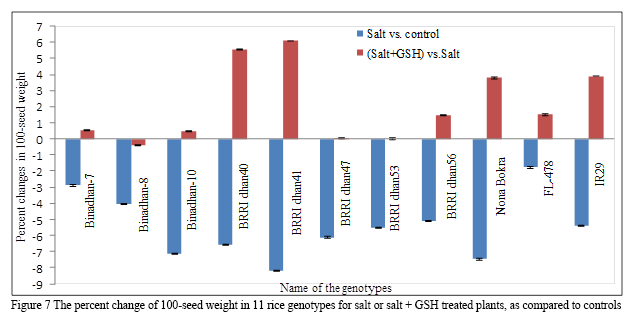 Figure 7 The percent change of 100-seed weight in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
1.2.9 Yield per panicle (g)
The genotype FL-478 showed the highest yield (3.12g) under control conditions, whereas the lowest yield (1.54 g) was found for IR29 under salt stress conditions (Table 2b). Imposition of salt stress resulted in a significant decrease in yield, the highest reduction (49.64%) was recorded for genotype IR29 followed by Nona Bokra (37.81%), BRRI dhan40 (34.56%), BRRI dhan41 (33.46%), BRRI dhan53 (31.43%), Binadhan-10 (30.17%), BRRI dhan56 (27.72%), FL-478 (23.32%), Binadhan-7 (20.57%), BRRI dhan47 (18.21%), and 15.07% for Binadhan-8 (Figure 8). Exogenous GSH treatment was found to increase yields, with the greatest increase (24.60%) was recorded for BRRI dhan40, followed by Nona Bokra (23.46%), IR29 (21.66%), BRRI dhan41 (18.49%), BRRI dhan53 (17.22%), BRRI dhan56 (16.35%), Binadhan-10 (11.94%), Binadhan-7 (10.88%), FL-478 (10.85%), Binadhan-8 (8.42%), with 3.51% for BRRI dhan47 (Table 3; Figure 8).
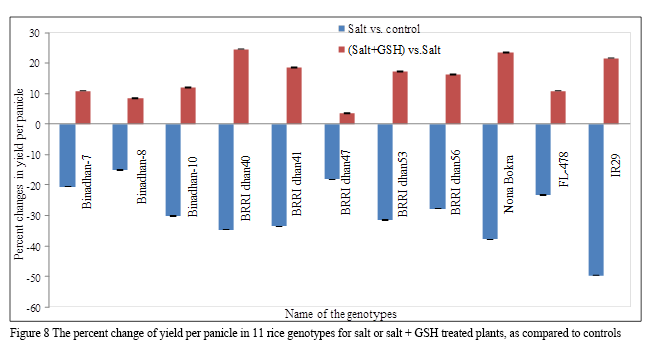 Figure 8 The percent change of yield per panicle in 11 rice genotypes for salt or salt + GSH treated plants, as compared to controls |
2 Discussion
Production of plants with agronomically and economically superior traits is the ultimate goal of a plant breeding programme. In the present study, the effects of salt stress on yield and yield contributing traits and the beneficial roles of exogenous GSH in promoting salt stress tolerance in rice was investigated. Many studies have shown that GSH is a vital antioxidant component for both the ascorbate-glutathione and glyoxalase systems, with multiple roles for ROS and MG removal/detoxification, and also improves plant growth and physiological adaptation under stressful conditions (Foyer and Noctor, 2003; Srivalli and Khanna-Chopra, 2008; Mostofa et al., 2014; Nahar et al., 2015a, b). This study aimed to test the hypothesis that an exogenous GSH treatment might be beneficial to modulate salt stress tolerance. The results presented clearly demonstrated that exogenous GSH exerts protective roles under salt stress, but this protective role was genotype specific.
Days to maturity was not affected for most genotypes under salt stress with or without exogenous GSH application. However, significant variations (p< 0.01) among the genotypes were found for this trait due to the inherent genetic variability of the breeding lines used. The number of effective tillers per plant was severely affected by salinity. Salinity is known to reduce the growth of plants at both the vegetative and reproductive stages (Rad et al., 2011), therefore, the reduced number of effective tillers per plant could be due to the negative consequence of salt stress during both stages of development. A reduction in the number of effective tillers per plant due to salinity has also been found in other studies (Baloch et al., 2003; Islam et al., 2007; Hasanuzzaman et al., 2009; Hussain et al., 2012; Joseph and Mohanan, 2013; Zaman et al., 2015). Although the number of effective tillers per plants in majority of the genotypes used in this study decreased under salinity, this negative impact was less in salt tolerant genotypes than in salt sensitive genotypes (Figure 1). Exogenous GSH application had a significant positive effect, increasing the number of effective tillers per plant under salt stress conditions (Table 2a; Table3). The positive influence of GSH was greater in the high yielding and salinity sensitive genotypes than for the salt tolerant genotypes (Figure 1).
Plant height for most rice genotypes was significantly affected by salt stress, except Binadhan-10 (Table 2a; Table3). Similar decreases in plant height in response to salt stress have been reported for plants, including rice (Thirumeni et al., 2001; Cristo et al., 2001; Islam et al., 2007; Hasanuzzaman et al., 2009; Hussain et al., 2012; Joseph and Mohanan, 2013; Sultana et al., 2014; Zaman et al., 2015). The results presented here and other research also indicates that the effect of salinity on plant growth was highly genetically determined (Hasanuzzaman et al., 2009). However, different physiological process involved for the reduction of plant height e.g. the suppression of growth are most likely due to osmotic imbalance, excess concentrations of Na+ and Cl- ions and ROS accumulation (Pareek et al., 1998; Bose et al., 2014). Importantly, application of exogenous GSH was found to improve plant growth under salt stress for many of the genotypes tested in this study (Table 2a; Table3; Figure 2), a trend also observed by other researchers (Cao et al., 2013; Teh et al., 2015).
Panicle length is also an important character that is directly related to grain yield. Panicle length was severely affected by salinity in this study (Figure 3) and similar reductions in panicle length, due to salinity, have been found by other researchers (Islam et al., 2007; Hasanuzzaman et al., 2009; Mahmood et al., 2009; Sultana et al., 2014; Rahman et al., 2015). Application of GSH to salt treated rice plants significantly increases panicle length as compared to salt treated plants only. In the present study, the highest positive response of GSH with respect to panicle length was found for the salt tolerant genotype Nona Bokra and lowest positive response to GSH was found for Binadhan-10 (Figure 3). In addition, the number of filled grains per panicle was severely affected by salinity in the genotypes tested. The greatest reduction in the number of filled grains per panicle was observed in the salt sensitive genotype and the least reduction in salt tolerant genotype (Table 2b; Table3; Figure 4). The reduction in the number of filled grains per panicle due to salinity has also been reported in other studies (Islam et al., 2007; Hasanuzzaman et al., 2009; Mahmood et al., 2009; Hussain et al., 2012; Joseph and Mohanan, 2013; Puteh and Mondal, 2014; Sultana et al., 2014; Zaman et al., 2015, Rahman et al., 2015). Grain filling in rice is highly sensitive to salinity stress and this cause poor seed set and grain yields (Aref and Rad, 2012). Increased sterility in panicle could be due to increased accumulation of Na+, and less K+, in plants under salinity stress. Another reason for failure of seed set could be due to the reduced availability of carbohydrates in developing spikelets (Abdullah et al., 2001). Exogenous GSH application significantly increased the number of filled grains per panicle in plants subjected to salt stress. This could be due to improved ROS scavenging and MG detoxification (Selote and Khanna-Chopra, 2004; Hossain and Fujita, 2009, 2013; Hossain et al., 2013c; Bae et al., 2013; Cao et al., 2015; Cevik and Unyayar, 2015).
The number of unfilled grains per panicle is one of the most important undesirable characters that has significant negative effects on crop yields. In the present study, the most unfilled grains per panicle were observed in the salt susceptible genotypes, whereas the least unfilled grains per panicle were observed in the salt tolerant genotypes (Table 2b; Table3; Figure 5). The increased number of unfilled grains per panicle due to salt stress has also been reported in other studies (Islam et al., 2007; Hasanuzzaman et al., 2009; Hussain et al., 2012; Joseph and Mohanan, 2013; Puteh and Mondal, 2014; Sultana et al., 2014; Zaman et al., 2015, Rahman et al., 2015). The reduction in seed set observed, could have been due to failure of stigma receptivity (Abdullah & Khan, 1997; Khatun & Flowers, 1995a, 1955b). Application of GSH to salt treated plants significantly reduced the number of unfilled grains per panicle for most of the genotypes tested, under salt stress (Figure 5).
Fertile grain is an important contributory factor to grain yield. Spikelet fertility was severely affected by salinity (Table 2b; Table3). The greatest reduction in spikelet fertility was observed in the salt sensitive genotypes and the least reduction was found in the salt tolerant genotypes (Figure 6). A reduction in spikelet fertility due to salinity was also found by other researchers (Islam et al., 2007; Mahmood et al., 2009; Puteh and Mondal, 2014; Sultana et al., 2014; Zaman et al., 2015, Rahman et al., 2015) and could have been due to a failure of grain formation caused by lack of viable pollen and lower seed set (Hasanuzzaman et al., 2009). Murty and Murty (1982) suggested that, the severe inhibitory effects of salts on fertility could be due to differential competition for carbohydrate supply between the growth of the vegetative parts of the plant and the developing panicles. Stigma receptivity and grain filling in rice are the most sensitive steps associated with grain development and seed set under saline conditions. Application of exogenous GSH significantly increased spikelet fertility of all genotypes of plants treated with salt. The highest and lowest increase of spikelet fertility percentage was observed in the salt sensitive and salt tolerant genotypes, respectively (Figure 6). Recently, it has been reported that GSH is essential for pollen germination in vitro and that GSH depletion in pollen grains triggered disturbances in auxin metabolism that could lead to inhibition of pollen germination (Zechmann et al., 2011). The positive role of GSH in improving pollen and spikelet fertility have also found in rice under drought stress (Roy, 2015). Selote and Khanna-Chopra (2004) also reported that heat stress induced pollen/spikelet sterility was associated with inefficient antioxidant defense systems and lower GSH and AsA levels in susceptible genotypes as compared to the tolerant genotypes. Therefore, the greater response of the sensitive genotype to GSH was probably due to depletion of endogenous GSH, however, further studies are needed to confirm this.
100-seed weight was severely affected by salinity (Table 2b; Table3; Figure 7) and this has also found by other researchers (Islam et al., 2007; Mahmood et al., 2009; Puteh and Mondal, 2014; Sultana et al., 2014; Zaman et al., 2015, Rahman et al., 2015). This might be due to lower accumulation of carbohydrates and other food materials due to salt stress (Hasanuzzaman et al., 2009). Application of GSH significantly increased 100-seed weight in salt treated plants of most of the rice genotypes, except Binadhan-8 and BRRI dhan53 (Figure 7). The genotypes Binadhan-8 and BRRI dhan53 are salt tolerant in nature and these genotypes probably contain the higher level of endogenous GSH (Selote and Khanna-Chopra, 2004). Transgenic plants over-expressing GSH biosynthetic genes also showed significantly higher 1000 seed weight under salt stress conditions (Choe et al., 2013).
Yield per panicle was severely affected by salinity, with the greatest reduction in yield per panicle found in salt sensitive genotypes and the least reduction was found in the salt tolerant genotypes (Table 2b; 3; Figure 8). The reduction of grain yield per panicle in rice in response to salt stress has also been reported in other studies (Islam et al., 2007; Hasanuzzaman et al., 2009; Mahmood et al., 2009; Hussain et al., 2012; Joseph and Mohanan, 2013; Puteh and Mondal, 2014; Sultana et al., 2014; Zaman et al., 2015, Rahman et al., 2015). Importantly, the application of GSH to salt affected plants significantly increase yields per panicle, compared to plants only exposed to salt, with the positive influence of GSH greater for the sensitive genotypes (Figure 8). Recent studies have also demonstrated the beneficial roles of exogenous GSH in modulating abiotic stress tolerance through the regulation of multiple stress responsive pathways and by enhancing ROS and MG detoxification systems (Qui et al., 2013; Cao et al., 2013; Mostofa et al., 2014; Cao et al., 2015; Teh et al., 2015; Nahar et al., 2015a, b, c). Transgenic plants over-expressing a GSH biosynthetic gene have also showed significantly higher grain yields under salt stress (Choe et al., 2013).
Based upon the above findings it can be concluded that the salt stress significantly limits the yield potential of many rice genotypes by inhibiting yield attributing traits. Exogenous GSH was found to modulate the salt stress tolerance for most of the rice genotypes tested, but the positive effects of exogenous GSH were greater in sensitive genotypes than for tolerant genotypes. A few exceptional responses were also demonstrated by the tolerant genotypes. The low response to exogenous GSH found for resistant genotypes was probably due to higher endogenous GSH or AsA contents or other inherent physiological and biochemical advantages of these genotypes. Therefore, the results of the present study warrants further research at the molecular level to find the answer of the question how exogenous GSH treatments improved salt stress tolerance. Moreover, further studies including investigating ROS and MG metabolism in both seedlings and plants at the reproductive stage, including pollen fertility/sterility studies, will give us further insights about the molecular mechanisms of salt stress tolerance as well as the functional role of exogenous GSH in modulating salt stress tolerance.
3 Materials and Methods
3.1 Plant materials and seedling growth
The experiment consisted of 11 diverse rice (Oryza sativa L.) genotypes including one salt tolerant land race (Nona Bokra), one salt tolerant advanced breeding line (FL-478), one salt sensitive variety (IR29), two salt non-tolerant variety (Binadhan-7, BRRI dhan56) and six moderately salt tolerant varieties (viz., Binadhan-8, Binadhan-10, BRRI dhan40, BRRI dhan41, BRRI dhan47, BRRI dhan53). Seedlings were grown in a seed-bed and one month old seedlings were transplanted to earthen pots. Two seedlings were maintained in each pot.
3.2 Experimental design
The experiment was carried out with four replications and three treatments (viz., Control, salt (200 mM NaCl) and salt+ 2mM GSH) following a Randomized Complete Block Design (RCBD) design.
3.3 Imposition of salt stress and application of exogenous GSH
Just before the emergence of panicle from flag leaf, rice plants were subjected to salt stress (200 mM NaCl) by giving salt (NaCl) solution with irrigation water. Two plants per pot were treated with 100 mM NaCl on alternate day to reach a final concentration of 200 mM NaCl. After 24 hour of first salt treatment, seedlings of each pot were sprayed with GSH solution (200ml of 2 mM GSH (Sigma-Aldrich, Germany) solution containing 0.001% Tween-20) using a hand sprayer. The glutathione solution was sprayed onto plants two times a day on every alternate day of the salt stress treatment. The same amount of distilled water was sprayed onto control plants.
3.4 Data collection of different parameters of growth and yield
Data on different yield and yield attributing traits, such as days to maturity, number of effective tillers per plant, plant height, panicle length, number of filled grains per panicle, number of unfilled grains per panicle, spikelet fertility, 100-seed weight and yield per panicle were collected.
3.5 Statistical analysis
Data recorded for different parameters were compiled and tabulated in proper form and statistical analysis were done by MSTAT-C considering RCBD design with two factors to find out the statistical significance of the experimental results. The data obtained were analysed following a two-way analysis of variance (ANOVA), genotype and treatment interaction and mean differences were compared by a least significant difference (LSD) test by using MSTAT-C.
Author’s contributions
Mohammad Anwar Hossain conceived the idea. B. M. Nahid Hussain, Mohammad Anwar Hossain and Sharif-Ar-Raffi participated in designing of the experiment. B. M. Nahid Hussain and Soleh Akram conducted the experiment and collected the data. B. M. Nahid Hussain and Sharif-Ar-Raffi analysed the data. B. M. Nahid Hussain prepared the manuscript and Mohammad Anwar Hossain, Sharif-Ar-Raffi and David J. Burritt critically evaluated the results and the manuscript, and rewrote sections as required. All authors read and approved the final manuscript.
Abdullah Z., Khan M.A., and Flowers T.J., 2001, Causes of sterility and seed set in rice under salinity stress, Journal of Agronomy and Crop Science, 187:25-32.
http://dx.doi.org/10.1046/j.1439-037X.2001.00500.x
Abdullah Z., Khan M.A., and Flowers T.J., 2002, Causes of sterility in rice under salinity stress, Prospects for Saline Agriculture, 37: 177-187
http://dx.doi.org/10.1007/978-94-017-0067-2_19
Abdullah Z., and Khan M.A., 1997, Influence of environmental stresses on reproductive physiology of selected crops, DFID (UK) and PARC (Pakistan), Research Bulletin Printo Grafic Publisher, Islamabad, Pakistan, pp.1-14
Akbar M., Yabuno Y., and Nakao S., 1972, Breeding for saline resistant varieties of rice: variability for salt tolerance among some rice varieties, Japanese Journal of Breeding, 22: 277-284
http://dx.doi.org/10.1270/jsbbs1951.22.277
Aref F., and Rad H.E., 2012, Physiological characterization of rice under salinity stress during vegetative and reproductive stages, Indian Journal of Science and Technology, 5(4): 2578-2586
Bae M.J., Kim Y.S., Kim S., Choe Y.H., Lee E.J., Kim Y.H., Park H.M., and Yoon H.S., 2013, Transgenic rice overexpressing the Brassica juncea gamma-glutamylcysteine synthetase gene enhances tolerance to abiotic stress and improves grain yield under paddy field conditions, Molecular Breeding, 31: 931–945
http://dx.doi.org/10.1007/s11032-013-9846-8
Baloch A.W., Soomro A.M., Javed M.A., Bughio H.R., Alam S.M., Bughio M.S., Mohammed T., and Mastoi N.N., 2003, Induction of salt tolerance in rice through mutation breeding, Asian Journal of Plant Sciences., 2(3): 273-276
http://dx.doi.org/10.3923/ajps.2003.273.276
Bose J., Rodrigo-Moreno A., and Shabala S., 2014, ROS homeostasis in halophytes in the context of salinity stress tolerance, Journal of Experimental Botany, 65(5): 1241-1257
http://dx.doi.org/10.1093/jxb/ert430 PMid: 24368505
Cai Y., Cao F., Cheng W., Zhang G., and Wu F., 2011a, Modulation of exogenous glutathione in phytochelatins and photosynthetic performance against Cd stress in the two rice genotypes differing in Cd tolerance, Biological Trace Element Research, 143: 1159-1173
http://dx.doi.org/10.1007/s12011-010-8929-1 PMid: 21191821
Cai Y., Cao F., Wei K., Zhang G., and Wu F., 2011b, Genotypic dependent effect of exogenous glutathione on Cd-induced changes in proteins, ultrastructure and antioxidant defense enzymes in rice seedlings, Journal of Hazardous Materials, 192: 1056–1066
http://dx.doi.org/10.1016/j.jhazmat.2011.06.011 PMid: 21741764
Cao F., Liu L., Ibrahim W., Cai Y., and Wu F., 2013, Alleviating effects of exogenous glutathione, glycinebetaine, brassinosteroids and salicylic acid on cadmium toxicity in rice seedlings (Oryza Sativa), Agrotechnology, 2: 107
Cao F., Cai Y., Liu L., Zhang M., He X., Zhang G., and Wu F., 2015, Differences in photosynthesis, yield and grain cadmium accumulation as affected by exogenous cadmium and glutathione in the two rice genotypes, Plant Growth Regulation, 75: 715–723
http://dx.doi.org/10.1007/s10725-014-9973-1
Cevik S., and Unyayar S., 2015, The effects of exogenous application of ascorbate and glutathione on antioxidant system in cultivated Cicer arietinum and wild type C. reticulatum under drought stress, Journal of Natural and Applied Science, 19(1): 91-97
Chaves M.M., Flexas J., and Pinheiro C., 2009, Photosynthesis under drought and salt stress, regulation mechanisms from whole plant to cell, Annals of Botany, 103: 551–560
http://dx.doi.org/10.1093/aob/mcn125 PMid: 18662937 PMCid: PMC2707345
Chen J.H., Jiang H.W., Hsieh E.J., Chen H.Y., Chien C.T., Hsieh H.L., and Lin T.P., 2012, Drought and salt stress tolerance of an Arabidopsis glutathione S-transferase U17 knockout mutant are attributed to the combined effect of glutathione & abscisic acid, Plant Physiology, 158(1): 340-351
http://dx.doi.org/10.1104/pp.111.181875 PMid: 22095046 PMCid: PMC3252094
Choe Y.H., Kim Y.S., Kim I.S., Bae M.J., Lee E.J., Kim Y.H., Park H.M., and Yoon H.S., 2013, Homologous expression of gamma-glutamylcysteine synthetase increases grain yield and tolerance of transgenic rice plants to environmental stresses, Journal of Plant Physiology, 170: 610-618
http://dx.doi.org/10.1016/j.jplph.2012.12.002 PMid: 23294545
Cristo E., Gonzalez M.C., Cardenas R.M., and Perez N., 2001, Salinity tolerance evaluation at the young stage of three new rice (Oryza sativa L.) lines by morpho-agronomic markers, Cultivos Tropicales, 22(2): 43-45
Dasgupta S., Hossain M.M., Huq M., and Wheeler D., 2014, Climate change, soil salinity, and the economics of high-yield rice production in coastal Bangladesh, Policy Research working paper; no. WPS 7140
http://dx.doi.org/10.1596/1813-9450-7140
El-Shabrawi H., Kumar B., Kaul T., Reddy M.K., Singla-Pareek S.L., and Sopory S.K., 2010, Redox homeostasis, antioxidant defense and methyl glyoxal detoxification as markers for salt tolerance in Pokkali rice, Protoplasma, 245: 85-96
http://dx.doi.org/10.1007/s00709-010-0144-6 PMid: 20419461
Foyer C.H., and Noctor G., 2003, Redox sensing and signaling associated with reactive oxygen in chloroplants, peroxisomes and mitochondria, Physiologia Plantarum, 119: 355-364
http://dx.doi.org/10.1034/j.1399-3054.2003.00223.x
Ghosh A., Pareek A., Sopory S.K., and Singla-Pareek S.L., 2014, A glutathione responsive rice glyoxalase II, OsGLYII-2, functions in salinity adaptation by maintaining better photosynthesis efficiency and anti-oxidant pool, The Plant Journal, 80: 93-105
http://dx.doi.org/10.1111/tpj.12621 PMid: 25039836
Hossain M.A., and Fujita M., 2013, Hydrogen peroxide priming stimulates drought tolerance in mustard (Brassica juncea L.), Plant Gene and Trait, 4: 109-123
Hossain, M.A., and Fujita M., 2009, Purification of glyoxalase I from onion bulbs and molecular cloning of its cDNA, Bioscience Biotechnology and Biochemistry, 73: 2007-2013
http://dx.doi.org/10.1271/bbb.90194 PMid: 19734676
Hasanuzzaman M., Fujita M., Islam M.N., Ahamed K.U., and Nahar K., 2009, Performance of four irrigated rice varieties under different levels of salinity stress, International Journal of Integrative Biology, 6: 85–90
Hossain M.A., Hossain M.Z., and Fujita M., 2009, Stress-induced changes of methylglyoxal level and glyoxalase I activity in pumpkin seedlings and cDNA cloning of glyoxalase I gene, Australian Journal of Crop Science, 3: 53-64
Hossain M.A., Mostofa M.G., and Fujita M., 2013a, Cross protection by cold-shock to salinity and drought stress-induced oxidative stress in mustard (Brassica campestris L.) seedlings, Molecular Plant Breeding, 4: 50-70
http://dx.doi.org/10.5376/mpb.2013.04.0007
Hossain M.A., Mostafa M.G., and Fujita M., 2013b, Heat-shock positively modulates oxidative protection of salt and drought stressed mustard (Brassica campestris L.) seedlings, Journal of Plant Science & Molecular Breeding, 2: 1-14
Hossain M.A., Ismail M.R., Uddin M.K., Islam M.Z., and Ashrafuzzaman M., 2013c, Efficacy of ascorbate-glutathione cycle for scavenging H2O2 in two contrasting rice genotypes during salinity stress, Australian Journal of Plant Science, 7(12): 1801-1808
Hossain H., Rahman M.A., Alam M.S., and Singh R.K., 2015, Mapping of quantitative trait loci associated with reproductive stage salt tolerance in rice, Journal of Agronomy and crop science, 201(1): 17-31
http://dx.doi.org/10.1111/jac.12086
Hoque A., Uraji M., Banu M.N., Mori I.C., Nakamura Y., and Murata Y., 2010, The effects of methylglyoxal on glutathione S-transferase from Nicotiana tabacum, Bioscience Biotechnology and Biochemistry, 74: 2124–2126
http://dx.doi.org/10.1271/bbb.100393 PMid: 20944411
Hussain M., Jang K.H., Farooq M., and Lee D.J., 2012, Morphological and physiological evaluation of Korean rice genotypes for salt resistance, International Journal of Agriculture and Biology, 14: 970‒974
Islam M.Z., Baset M.A., Islam M.R., and Akter A., 2007, Effect of different saline levels on growth and yield attributes of mutant rice, Journal of Soil and Nature, 1(2): 18-22
Joseph E.A., and Mohanan K.V., 2013, A study on the effect of salinity stress on the growth and yield of some native rice cultivars of Kerala state of India, Agriculture, Forestry and Fisheries, 2(3): 141-150
http://dx.doi.org/10.11648/j.aff.20130203.14
Kader M.A., and Lindberg S., 2010, Cytosolic calcium and pH signalling in plants under salinity stress, Plant Signalling & Behavior, 5: 233-238
http://dx.doi.org/10.4161/psb.5.3.10740 PMid: 20037468 PMCid: PMC2881266
Karim Z., Hussain S.G., and Ahmed M., 1990, Salinity problems and crop intensification in the coastal regions of Bangladesh, BARC soil publication no.33, Dhaka, pp.63
Khatun S., and Flowers T.J., 1995a, The estimation of pollen viability in rice, Journal of Experimental Botany, 46: 151-154
http://dx.doi.org/10.1093/jxb/46.1.151
Khatun S., and Flowers T.J., 1995b, Effect of salinity on seed set in rice, Plant Cell Environ., 18: 61-67
http://dx.doi.org/10.1111/j.1365-3040.1995.tb00544.x
Labudda M., and Azam F.M.S., 2014, Glutathione-dependent responses of plants to drought: a review, Acta Societatis Botanicorum Poloniae, 83: 3–12
http://dx.doi.org/10.5586/asbp.2014.003
Lauchli A., and Grattan S.R., 2007, Plant growth and development under salinity stress. In: Jenks M.A., Hasegawa P.M., and Jain M.S., (eds.), Advances in molecular breeding toward drought and salt tolerant crops, Springer, Dordrecht, Netherlands, pp.1–32
http://dx.doi.org/10.1007/978-1-4020-5578-2_1
Mahmood A., Latif T., and Khan M.A., 2009, Effect of salinity on growth, yield and yield components in basmati rice germplasm, Pakistan Journal of Botany, 41(6): 303-304
Malik A.R., Zahida H.P., and Muhammad S.M., 2008, Genetic diversity analysis of traditional and improved cultivars of Pakistani rice (Oryza sativa L.) using RAPD markers, Electronic Journal of Biotechnology, 11(3): 1-10
Maas E.V., and Grattan S.R., 1999, Crop yields as affected by salinity, In: Skaggs R. W., and J. van Schilfgaarde (ed.), Agricultural drainage, American Society of Agronomy, Madison, USA, pp.55–108
Meloni D.A., Oliva M.A., Martinez C.A., and Cambraia J., 2003, Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress, Environmental and Experimental Botany, 49: 69–76
http://dx.doi.org/10.1016/S0098-8472(02)00058-8
Mostofa M.G., Seraj Z.I., and Fujita M., 2014, Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings, Protoplasma, 251: 1373–1386
http://dx.doi.org/10.1007/s00709-014-0639-7 PMid: 24752795
Mostofa M.G., Saegusa D., Fujita M., and Tran L.S.P., 2015a, Hydrogen sulfide regulates salt tolerance in rice by maintaining Na+/K+ balance, mineral homeostasis and oxidative metabolism under excessive salt stress, Frontiers Plant Science, 6: 1055
http://dx.doi.org/10.3389/fpls.2015.01055 PMid: 26734015 PMCid: PMC4685665
Mostofa M.G., Hossain M.A., and Fujita M., 2015b, Trehalose pretreatment induces salt tolerance in rice seedlings: oxidative damage and co-induction of antioxidant defense and glyoxalase systems, Protoplasma, 252(2): 461-475
http://dx.doi.org/10.1007/s00709-014-0691-3PMid: 25164029
Murty P.S.S., and Murty K.S., 1982, Spikelet sterility in relation to nitrogen and carbohydrate content in rice, Indian Journal of Plant Physiology, 25: 40-48
Nahar K., Hasanuzzaman M., Alam M.M., and Fujita M., 2105a, Glutathione-induced drought stress tolerance in mung bean: coordinated roles of the antioxidant defense and methylglyoxal detoxification systems, AoB Plants, 7: plv069
http://dx.doi.org/10.1093/aobpla/plv069 PMid: 26134121 PMCid: PMC4526754
Nahar K., Hasanuzzaman M., Alam M.M., and Fujita M., 2015b, Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiate L.) by modulating antioxidant defense and methylglyoxal detoxification system, Environmental and Experimental Botany, 112: 44–54
http://dx.doi.org/10.1016/j.envexpbot.2014.12.001
Nahar K., Hasanuzzaman M., Alam M.M., and Fujita M., 2015c, Roles of exogenous glutathione in antioxidant defense system and methylglyoxal detoxification during salt stress in mung bean, Biologia. Plantarum, 59(4): 745-756
http://dx.doi.org/10.1007/s10535-015-0542-x
Pareek A., Singla S.L., and Grover A., 1998, Proteins alterations associated with salinity, desiccation, high and low temperature stresses and abscisic acid application in seedlings of Pusa 169, a high yielding rice (Oryza sativa L.) cultivar, Current Science, 75(10): 1023-1035
Puteh A.B., and Mondal M.M.A., 2014, Salinity stress during booting and heading stages affects yield in rice, Life Science Journal, 11(6): 223-226
Qui B.Y., Zeng F.R., Cai S.G., Wu X.J., Haider S.I., Wu F.B., and Zhang G.P., 2013, Alleviation of chromium toxicity in rice seedlings by applying exogenous glutathione, Journal of Plant Physiology, 170: 772-779
http://dx.doi.org/10.1016/j.jplph.2013.01.016 PMid: 23523466
Ramírez L., Bartoli C.G., and Lamattina L., 2013, Glutathione and ascorbic acid protect Arabidopsis plants against detrimental effects of iron deficiency, Journal of Experimental Botany, 64(11): 3169-3178
http://dx.doi.org/10.1093/jxb/ert153 PMid: 23788722
Rad H.E., Aref F., Khaledian M., Rezaei M., Amiri E., and Falakdehy O.Y., 2011, The effects of salinity at different growth stages of rice yield, ICID 21st International Congress on Irrigation and Drainage, New Delhi, India, pp.155-165
Rahman M.S., Haque M.A., and Islam M.T., 2015, Salinity affects flag leaf chlorophyll and yield attributes of rice genotypes, Journal of Bioscience and Agriculture Research, 4(2): 80-85
Roy K., 2015, Effects of exogenous glutathione in drought stress tolerance of rice varieties, Thesis for M.S., Bangladesh Agricultural University, Mymensingh
Sairam R.K., Rao V.K., and Srivastava G.C., 2002, Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration, Plant Science, 163: 1037–1046
http://dx.doi.org/10.1016/S0168-9452(02)00278-9
Selote D.S., and Khanna-Chopra R., 2004, Drought-induced spikelet sterility is associated with an inefficient antioxidant defense in rice panicles, Physiologia Plantarum, 121: 462–471
http://dx.doi.org/10.1111/j.1399-3054.2004.00341.x
Srivalli S., and Khanna-Chopra R., 2008, Role of glutathione in abiotic stress tolerance, In: Khan N.A., Singh S., Umar S., (ed.), Sulfur assimilation and abiotic stress in plants, Springer, Berlin, pp.207-225
http://dx.doi.org/10.1007/978-3-540-76326-0_10
Sultana T., Islam R., Chowdhury M.S.N., Islam M.S., Hossain M.E., and Islam M.M., 2014, Performance evaluation of two rice varieties under different levels of NaCl salinity stress, Bangladesh Research Publication Journals, 10(2): 186-195
Teh C.Y., Mahmood M., Shaharuddin N.A., and Ho C.L., 2015, In vitro rice shoot apices as simple model to study the effect of NaCl and the potential of exogenous proline and glutathione in mitigating salinity stress, Plant Growth Regulation, 75: 771–781
http://dx.doi.org/10.1007/s10725-014-9980-2
Thirumeni S., Anuratha A., Ramanadane T., and Paramasivam K., 2001, Effect of salinity on seed germination and seedling growth of rice varieties, Crop Researh-Hisar, 22(3): 335-338
Uddin M.N., 2005, Changes of environmental parameters due to salinity intrusion in the southern region of Bangladesh, Thesis for M.S., Institute of water and Flood management, BUET, Dhaka
Upadhyaya C.P., Venkatesh J., Gururani M.A., Asnin L., Sharma K., Ajappala H., and Park S.W., 2011, Transgenic potato overproducing L-ascorbic acid resisted an increase in methylglyoxal under salinity stress via maintaining higher reduced glutathione level and glyoxalase enzyme activity, Biotechnology Letters, 33: 2297-2307
http://dx.doi.org/10.1007/s10529-011-0684-7 PMid: 21750996
Wang R., Liu S., Zhou F., and Hua C., 2014, Exogenous ascorbic acid and glutathione alleviate oxidative stress induced by salt stress in the Chloroplasts of Oryza sativa L., Zeitschrift für Naturforschung C, 69(5-6): 226-236
http://dx.doi.org/10.5560/znc.2013-0117 PMid: 25069161
Yamne K., Taniguchi M., and Miyake H., 2012, Salinity induced subcellular accumulation of H2O2 in leaves of rice, Protoplasma, 249: 301-308
http://dx.doi.org/10.1007/s00709-011-0280-7 PMid: 21533665
Zaman K.A., Halim M.A., Hossain M.A., Razzaque M.A., Zamil S.S., and Khatun F., 2015, Consequences of salinity on growth and yield performance of rice (Oryza sativa L.), International Journal of Business, Social and Scientific Research, 3(4): 223-229
Zeng L., and Shannon M.C., 2000, Salinity effects on seedling growth and yield components of rice, Crop Science, 40: 996–1003
http://dx.doi.org/10.2135/cropsci2000.404996x
Zechmann B., Koffler B.E., and Russell S., 2011, Glutathione synthesis is essential for pollen germination in vitro, BMC Plant Biology 11: 54
http://dx.doi.org/10.1186/1471-2229-11-54 PMid: 21439079 PMCid: PMC3078877



. PDF(730KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. B. M. Nahid Hussain
. Soleh Akram
. Sharif-Ar- Raffi
. David J. Burritt
. Mohammad Anwar Hossain
Related articles
. Salt stress
. Exogenous glutathione
. Reproductive stage
. Stress tolerance
. Rice ( Oryza sativa L.)
Tools
. Email to a friend
. Post a comment