Molecular Cloning of Promoter of AP3 Gene from Arabidopsis thaliana Ecotype (Col) and Construction of the Plant Expression Vector 
2. College of Forestry, Henan University of Science and Technology, Henan, 471003, P.R. China
 Author
Author  Correspondence author
Correspondence author
Plant Gene and Trait, 2012, Vol. 3, No. 5 doi: 10.5376/pgt.2012.03.0005
Received: 30 Jan., 2012 Accepted: 24 Feb., 2012 Published: 01 Mar., 2012
Fan et al., 2012, Molecular Cloning of Promoter of AP3 Gene from Arabidopsis thaliana Ecotype (Col) and Construction of the Plant Expression Vector, Plant Gene and Trait, Vol.3, No.5 22-27 (doi: 10.5376/pgt.2012.03.0005)
Belonging to MADS-Box gene family, APETALA3 (AP3) gene of Arabidopsis floral organ class B is expressed in petals and stamens specifically. AP3 gene which encodes a transcription factor controls the development of petals and stamens of dicotyledons by cooperation with class A and class C genes, respectively. The results showed that the promoter of AP3 was the flower-specific expression promoter. Therefore, the cloning and functional identification of AP3 gene promoter from Arabidopsis thaliana will play significant effects on the oriented improvement of the commercial traits related to the flowers of ornamental plants. Here, a pair of specific PCR primers was designed according to the promoter sequence of AP3 gene of Arabidopsis thaliana ecotype (Ler) reported in GenBank (U30729). A length of 1 767 bp of AP3 gene promoter was obtained from Arabidopsis thaliana ecotype (Col) by using PCR technique with high-fidelity DNA polymerase KOD-plus, named as pAtAP3, the GenBank accession No. is FJ619533. Online Bl2seq analysis indicated that the sequence similarity between pAtAP3 and U30729 was 98%. It also showed that the sequence similarity between pAtAP3 and the BAC clone T12E18 (AL132971) of Arabidopsis thaliana ecotype (Col) from 9 264 to 11 030 was up to 100%, and its downstream sequence encoded AP3 protein (CAB81799), which approved that the sequence we cloned was the promoter of AP3 gene of Arabidopsis thaliana ecotype (Col). Online PLACE analysis displayed that pAtAP3 contained the basic cis-elements of promoter such as TATA-box and CAAT-box. It also comprised several cis-elements related to flower-specific expression, such as CArG1, CArG2, CArG3 and anther-box. Moreover, plant expression vector of pAtAP3::GUS was successfully constructed in this research, which would lay the foundation for functional identification of the AP3 promoter of Arabidopsis thaliana ecotype (Col).
Based on the study of floral homeotic mutants of Arabidopsis thaliana and Antirrhinum majus L., Coen and Meyerowitz (1991) proposed the ABC model of flower development, subsequently, it was extended to the ABCD model later by Angenent and Colombo (1996). In the genetic network of flower development, flower organ identity genes determine floral organ primordia developed into sepals, petals, stamens or carpels. The class B flower organ identity gene, APETALA3 (AP3), encoding transcription factors, belongs to MADS-Box gene family. Specifically expressed in petal and stamen, AP3 gene synergistic controlled the petal and stamen development of dicotyledons with class A and class C identity genes (Krizek and Meyerowits, 1996). Promoter expression analysis displayed that AP3 promoter was the flower-specific expression promoter (Verdonk et al., 2008).
When using biotechnology for genetic improvement on important commercial traits in plant, if the heterologous gene expression was driven by constitutive promoter such as CaMV 35S, it may inhibit the growth and development of the transgenic plants. Under the control of one flower-specific expression promoter, the foreign gene will be specifically expressed in floral organs, which will increase the regional expression, and will not have an adverse development effects on transgenic plants. Therefore, the flower-specific expression promoters have an important application potential to directionally improve the comercial traits of landscape plants (Fan et al., 2010; Gao et al., 2010). Currently, AP3 promoter of Arabidopsis ecotype (Ler) has been cloned (Koch et al., 2001), and site-specific mutation analysis revealed that cis-elements such as CArG1, CArG2 and CArG3 are closely related to flower-specific of AP3 expression (Tilly et al., 1998). Driven by Ap3 promoter from Ler ecotype Arabidopsis, the heterologous gene ipt from Agrobacterium tumefaciens can specifically expressed in the floral organ of transgenic petunia (Petunia hybrida), resulting in the significant increase of flower diameter and fresh weight, but almost same size of vegetative to that of non-transgenic plants (Verdonk et al., 2008). This study provides a theoretical and practical basis for further application of flower-specific expression promoter in improving the commercial floral traits of landscape plants. The three most useful ecotypes in Arabidopsis are the Landsberg erecta (Ler), Columbia (Col) and Wassilewskija (Ws), respectively, among these different ecotypes, there are large differences in morphological development and physiological response (Zhang et al., 2006). So far, AP3 promoter of Arabidopsis ecotype (Col) has not been reported, so we cloned the AP3 promoter from Arabidopsis ecotype (Col) (GenBank accession No. FJ619533), conducted bioinformatics analysis and constructed pAtAP3::GUS plant expression vector so as to lay ground to further explore the function and expression mechanism of AP3 promoter of Arabidopsis ecotype (Col) and accumulate information on the directed improvement of the commercial traits related to flower in landscape plants.
1 Results and Analysis
1.1 PCR amplification of pAtAP3
Taken total DNA extracted from young leaves of Arabidopsis ecotype (Col) as DNA template, we successfully amplified a fragment of AP3 gene promoter, pAtAP3, with high-fidelity KOD-plus DNA polymerase. The product was about 1 800 bp, which was consistent with the predicted length (Figure 1).
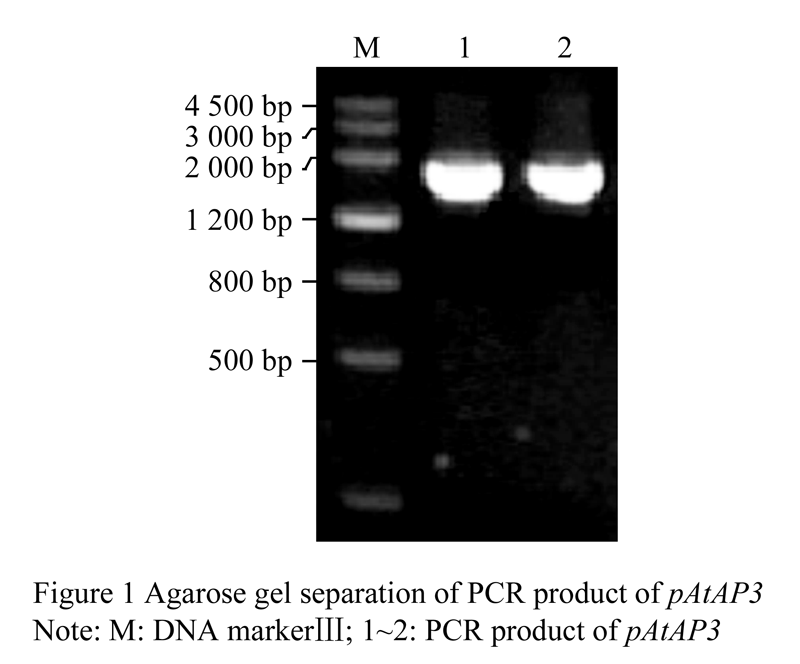 Figure 1 Agarose gel separation of PCR product of pAtAP3 |
1.2 Cloning of pAtAP3
After recovered with DNA purification Kit, the target PCR products were added nucleotide A at 3' end. Then they were ligated to pMD18-T overnight and the ligated products were transformed into competent cells of E. coli DH5α by heat shock method and incubated at 37℃ overnight. The transformants were firstly selected by colony PCR and plasmid PCR, and then were identified by double enzyme digestion with Hindâ…¢ and Xbaâ… . The digest products of the positive recombinants, 1 800 bp and 2 800 bp (Figure 2), were consistent with the predicted length, which indicated that they were suitable to be sequenced.
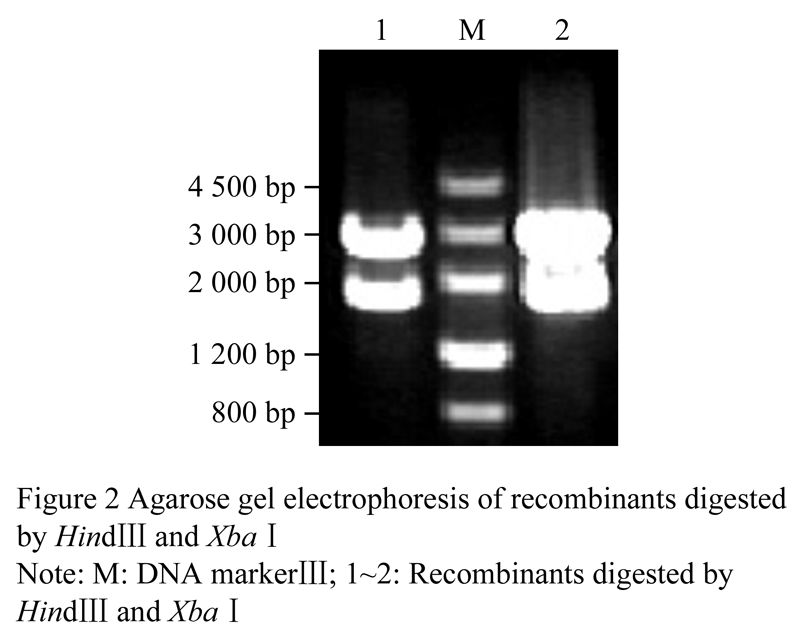 Figure 2 Agarose gel electrophoresis of recombinants digested by Hindâ…¢ and Xbaâ… |
1.3 Sequencing and sequence analysis of pAtAP3
The sequencing results of the two recombinants of pAtAP3 were the same and the length was 1 767 bp (Figure 3). Pairwise sequence alignment analysis based on bl2seq program showed that the sequence similarity between pAtAP3 and U30729 was up to 98%, indicating that AP3 promoter sequences between different ecotypes of Arabidopsis appeared polymorphism. Online BLAST analysis revealed that the sequence similarity between cloned pAtAP3 and BAC clone (AL132971) from 9 264 to 11 030 in No. 3 chromosome of Arabidopsis ecotype (Col) was up to 100%, and its downstream sequence of the cloned promoter exactly encoded APETALA3. Therefore, the cloned sequence was identified as the AP3 promoter of Arabidopsis ecotype (Col), and the GenBank accession No. was FJ619533. Online PLACE analysis showed that there was a TATA box (TTATTT) between nucleotide 1 205 bp to 1 210 bp, whose function was to ensure the accurate transcription start. It also contained 6 CAAT boxes, from 154 bp to 157 bp, 328 bp to 331 bp, 878 bp to 881 bp, 1 252 bp to 1 255 bp, 1 469 bp to 1 472 bp and 1 619 bp to 1 622 bp, respectively, which determined the start frequency of the promoter. Three conserved elements CArG-box binding to MADS-box protein in pAtAP3were directly related to the specific expression of AP3 gene in petals and stamens (Koch et al., 2001; Tilly et al., 1998). CArG1-box (GTTTACATAAATGGAAAA), located from 1 551 bp to 1 568 bp, was the binding sites of heterodimer AP3/PI. If the cis-element of CArG2-box (CTTACCTTTCATGGATTA) which was located from bp 1 599 to 1 616 bp was mutated, it can lead to lowwer expression of AP3 gene in petals, but without any changes in stamens. The 3rd CArG3-box (CTTTCCATTTTTAGTAAC) was located from 1 624 bp to 1 641 bp. Two anther box (AGAAA) located from 114 bp to 118 bp and from 955 bp to 959 bp, respectively, were the core cis-elements of AP3 promoter specifically expressed in stamen (Tunen et al., 1989). However, the G-box (Staiger et al., 1989) and TACPyAT-box elements (van der Meer et al., 1992) contained in promoter of CHS gene from Petunia were not detected in pAtAP3.
 Figure 3 Sequence of pAtAP3 |
1.4 Construction of plant expression vector pAtAP3::GUS
The empty vector pBI121 and recombinant pMD18T-pAtAP3 were all digested by Hindâ…¢ and Xba I. Then the large digestion product of pBI121 (Figure 4) and the small digestion fragment of pMD18-T-pAtAP3 (Figure 2) were recovered, respectively. The recovered products were ligated and transformed into E. coli DH5α. And then the colony PCR and plasmid PCR were carried out to characterize the correct construct of pAtAP3::GUS. The electrophoresis result showed that the 1 800 bp target fragment was amplified (Figure 5), which revealed that the plant expression vector pAtAP3::GUS was successfully constructed.
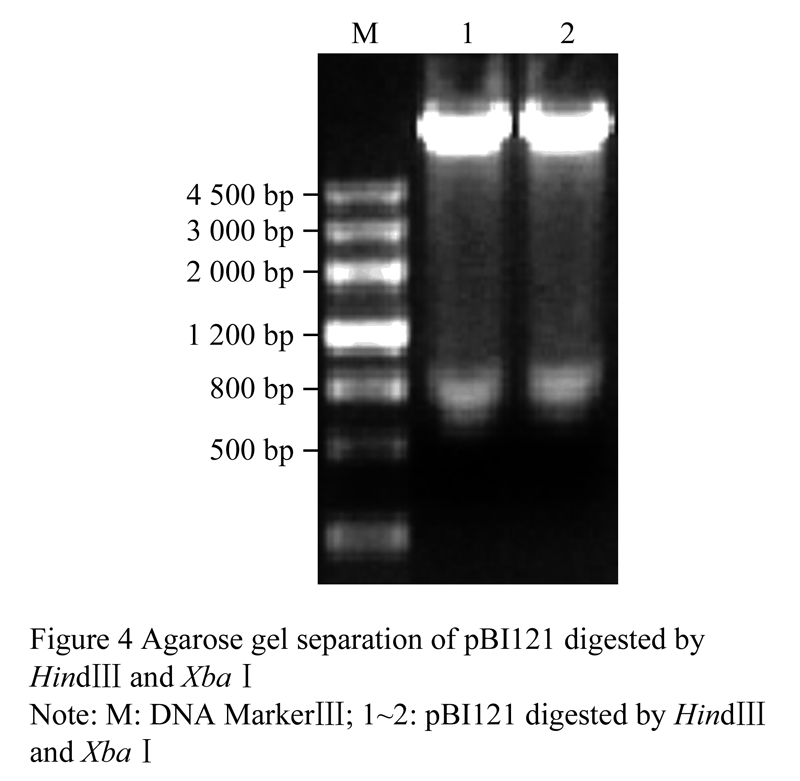 Figure 4 Agarose gel separation of pBI121 digested by HindIII and Xbaâ… sequence of pAtAP3 |
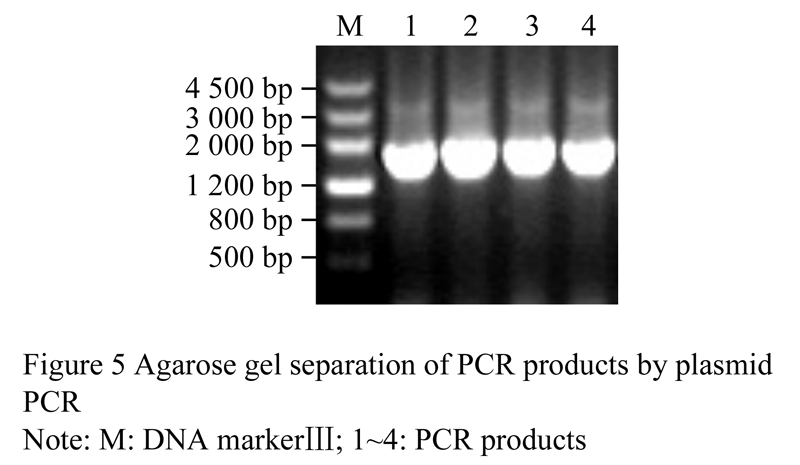 Figure 5 Agarose gel separation of PCR products by plasmid PCR |
2 Discussion
The genetic engineering of landscape plant was involved in improvement of lots of traits such as flower color, flower type and flower diameter, so cloning and identification of flower-specific expression promoter has some important application potential on oriented improvement of the commercial traits related to floral organs (Fan et al., 2010; Gao et al., 2010). Cucumber (Cucumis sativus L.) has served as a model plant to understand hormone regulation in unisexual flower development. It has been confirmed the role of ethylene in the promotion of female cucumber flowers. Under the control of AP3 promoter of Arabidopsis ecotype (Ler), cucumber gene CsACO2 was overexpressed in transgenic Arabidopsis. It showed that transgenic Arabidopsis displayed normal vegetative morphology, but all flowers were sterile, which was exactly caused by abnormal stamen. However, carpels retained normal function with no obvious morphological change. The abnormal stamen morphological change was resulted from increasing ACO2 enzyme activity about 20% in floral bud (Duan et al., 2008), which confirmed that the potential application of AP3 promoter in fine regulation of floral traits.
In this study, we cloned pAtAP3 from Arabidopsis ecotype (Col), which was assigned GenBank accession as FJ619533. Bioinformatic analysis showed that it contained several cis-elements related to flower-specific expression. And the plant expression vector pAtAP3:: GUS was successfully constructed. Therefore, it would provide a material to further explore the expression pattern and the mechanism of flowers-pecific expression of pAtAP3. It also would lay the foundation to apply pAtAP3 for directly improvement the commercial traits of landscape plant in the future.
3 Materials and Methods
3.1 Plant materials
Young rosette leaves of Arabidopsis thaliana ecotype Col were used to extract total DNA.
3.2 Strain, plasmid and reagent
KOD-plus DNA polymerase was purchased from TOYOBO Co. LTD. Plant expression vector pBI121 and Escherichia coli strain DH5α were kept in our own laboratory. Cloning vector pMD18-T, Agarose Gel DNA Purification Kit, MiniBEST Plasmid Purification Kit, restriction endonucleases Hindâ…¢ and Xbaâ… were from TaKaRa Biotechnology (Dalian) Co. LTD. DNA Marker III and Taq DNA polymerase were bought from Tiangen Biotech (Beijing) Co., LTD.
3.3 Total DNA extraction of Arabidopsis leaves and primer design
Total DNA was isolated from young Arabidopsis leaves according to modified CTAB method (Mummenhoff and Koch, 1994), and the final concentration of total DNA was adjusted to 50 ng/μL. Based on AP3 promoter sequence of Arabidopsis thaliana ecotype Ler reported in GenBank (U30729) and multiple cloning sites on the upstream and downstream of Cauliflower Mosaic Virus 35S promoter in pBI121 vector, a pair of specific PCR primers was designed with Primer Premier 5.0 software. Forward primer 5'-AAGCTTCTTAAGAATTATAGTAGCACTTGTT-3', containing self Hindâ…¢ restriction sites (underline), and the downstream primer 5'-TGCTCTAGAATTCTTCTCTCTTTGTTTA ATC-3', inserted with Xbaâ… restriction sites (underline), were synthesized by Beijing AuGCT Biotechnology Co., Ltd.
3.4 PCR amplification of pAtAP3
50 µL PCR reaction system was composed of 5 µL 10×PCR Buffer, 5 µL 2.0 mmol/L each dNTP, 1 µL total DNA, 3 µL 25 mmol/L MgSO4, 1.5 µL 10 pmol/µL forward and reverse primer, respectively, 1 µL KOD-plus DNA polymerase and 32 µL ddH2O. Thermocycling was performed at 95℃ for 4 min, then at 95℃ for 30 s, 52℃ for 30 s, 68℃ for 2 min 30 s for 35 cycles, 68℃ for 7 min and finally kept at 4℃. The PCR products were separated by 1% Agarose gel electrophoresis.
3.5 Cloning and sequencing of pAtAP3
The target PCR products were recovered with TaKaRaTM Agarose Gel DNA Purification Kit. Because nucleotide A can't be added to the 3' end of PCR products amplified by high-fidelity KOD-Plus DNA polymerase, it is necessary to add nucleotide A at its 3' end before the purified target fragment was cloned into pMD18-T vector. The adding nucleotide A reaction system was 10 μL, containing purified PCR products 8 μL, 2.5 mmol/L each dNTP 0.8 μL, PCR Buffer1 μL, and Taq DNA polymerase 0.2 μL. This reaction was carried out at 72℃ for 30 min. Then these products were ligated to pMD18-T with T4 DNA ligase and the ligated products were transformed into competent cells of E. coli DH5α by using heat shock method. The transformants were selected by colony PCR and plasmid PCR, and then were identified by double enzyme digestion of Hindâ…¢ and Xbaâ… . The positive recombinants were named as pMD18-T-pAtAP3, and were sequenced by Sangon Biotech (Shanghai) Co. Ltd.
3.6 Bioinformatic analysis of pAtAP3 sequence
The sequence of pAtAP3 was submitted to GenBank for accession number (http://www.ncbi.nlm.nih.gov/). Sequences of pAtAP3 and U30729 were aligned by online bl2seq program (http://blast.ncbi.nlm.nih.gov/ Blast.cgi). cis-elements of pAtAP3 were analyzed with online analysis tools PLACE (http://www.dna.affrc.go. jp/PLACE/signalscan.html) (Higo et al., 1999).
3.7 Construction of plant expression vector pAtAP3::GUS
In order to further explore the expression pattern of pAtAP3, it is necessary to construct plant expression vector pAtAP3::GUS. Firstly, the positive recombinant pMD18-T-pAtAP3 and empty vector pBI121 were digested by Hindâ…¢ and Xbaâ… . Then the small digestion fragment of recombinant pMD18-T-pAtAP3 and the large fragment digestion product of pBI121 were recovered, respectively. Later the purified products were ligated overnight, the ligated products were transformed into E. coli DH5α competent cells and cultured on LB medium with 50 µg/mL Kanamycin. The plant expression vector pAtAP3::GUS was identified by colony PCR and plasmid PCR.
Authors' Contributions
SPG, XGH and HWX conducted this experiment; GAS and XSK participated in the experiment design; FBY was the person who took charge of this project, including experiment design, data analysis, writing and modifying of the manuscript. All authors have read and approved the final manuscript.
Acknowledgements
This study was financially supported by the National Natural Science Foundation of China (30740013) and Program for young teachers in University and college of Henan Province (2010GGJS-075). Authors appreciate two anonymous reviewers for their useful critical comments and revising advice to this paper.
References
Angenent G.C., and Colombo L., 1996, Molecular control of ovule development, Trends in Plant Sci., 1(7): 228-232
Coen E.S., and Meyerowitz E.M., 1991, The war of the whorls: genetic interactions controlling flower development, Nature, 353(6339): 31-37
http://dx.doi.org/10.1038/353031a0 PMid:1715520
Duan Q.H., Wang D.H., Xu Z.H., and Bai S.H., 2008, Stamen development in Arabidopsis is arrested by organ-specific overexpression of a cucumber ethylene synthesis gene CsACO2, Planta, 228(4): 537-543
http://dx.doi.org/10.1007/s00425-008-0756-7 PMid:18506477
Fan B.Y., Gao S.P., Hou X.G., and Shi G.A., 2010, Cloning of a flower-specific expression promoter from Arabidopsis thaliana and its plant expression vector construction, Forestry Studies in China, 12(4): 201-205
http://dx.doi.org/10.1007/s11632-010-0408-4
Gao S.P., Fan B.Y., Shi G.A., Jia X.P., and Kong X.S., 2010, Cloning and sequence analysis of promoter of chalcone synthase gene from Petunia Hybrida and construction of plant expression vector, Northern Horticulture, 17: 155-158
Higo K., Ugawa Y., Iwamoto M., and Korenaga T., 1999, Plant cis-acting regulatory DNA elements (PLACE) database: 1999, Nucleic Acids Research, 27(1): 297-300
http://dx.doi.org/10.1093/nar/27.1.297 PMid:9847208 PMCid:148163
Koch M.A., Weisshaar B., Kroymann J., Haubold B., and Mitchell-Olds T., 2001, Comparative genomics and regulatory evolution: conservation and function of the Chs and Apetala3 promoters, Molecular Biology and Evolution, 18(10): 1882-1891
http://dx.doi.org/10.1093/oxfordjournals.molbev.a003729 PMid:11557794
Krizek B.A., and Meyerowits E.M., 1996, The Arabidopsis homeotic genes APETAGA3 and PISTILLATA are sufficient to provide the B class organ identity function, Development, 122(1): 11-22 PMid:8565821
Mummenhoff K., and Koch M., 1994, Chloroplast DNA restriction site variation and phylogenetic relationships in the genus Thlaspi sensu lato (Brassicaceae), Syst. Bot., 19(1): 73-88
http://dx.doi.org/10.2307/2419713
Staiger D., Kaulen H., and Schell J., 1989, A CACGTG motif of the Antirrhinum majus chalcone synthase promoter is recognized by an evolutionary conserved nuclear protein, Proc. Natl. Acad. Sci., USA, 86(18): 6930-6934
http://dx.doi.org/10.1073/pnas.86.18.6930
Tilly J.J., Allen D.W., and Jack T., 1998, The CArG boxes in the promoter of the Arabidopsis floral organ identity gene APETALA3 mediate diverse regulatory effects, Development, 125(9): 1647-1657 PMid:9521903
Tunen A.J., Hartman S.A., Mur L.A., and Mol J.N.M., 1989, Regulation of chalcone flavonone isomerase (CHI) gene expression in Petunia hybrida: the use of alternative promters in corolla, anthers and pollen, Plant Mol. Biol., 12(5): 539-551
http://dx.doi.org/10.1007/BF00036968r
van der Meer I.M., Brouwer M., Spelt C.E., Mol J.N.M., and Stuitje A.R., 1992, The TACPyAT repeats in the chalcone synthase promoter of Petunia hybrida act as a dominant negative cis-acting module in the control of organ-specific expression, The lant J., 2(4): 525-535
http://dx.doi.org/10.1046/j.1365-313X.1992.t01-23-00999.x PMid:1344889
Verdonk J.C., Shibuya K., Loucas H.M., Colquhoun T.A., Underwood B.A., and Clark D.G., 2008, Flower-specific expression of the Agrobacterium tumefaciens isopentenyltransferase gene results in radial expansion of floral organs in Petunia hybrida, Plant Biotechnology Journal, 6(7): 694-701
http://dx.doi.org/10.1111/j.1467-7652.2008.00349.x PMid:18482222
Zhang Z.Z., Xu Y.Q., and Huang H., 2006, Aribidopsis, a powerful tool for exploring the mysteries of plant kingdom, Chinese Bulletin of Life Sciences, 18(5): 442-446



. PDF(942KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Bingyou Fan
. Shuiping Gao
. Xiaogai Hou
. Huawei Xu
. Guoan Shi
. Xiangsheng Kong
Related articles
. Arabidopsis thaliana
. Ecotype Col
. Promoter of APETALA3
. Plant expression vector
Tools
. Email to a friend
. Post a comment