Research Article
Clonal Growth Characteristic of Epiphytic Orchid Dendrobium sinense 
2 Key Laboratory of Germplasm Resources of Tropical Special Ornamental Plants of Hainan Province, College of Forestry, Hainan University, Haikou, 570228, China
 Author
Author  Correspondence author
Correspondence author
Plant Gene and Trait, 2021, Vol. 12, No. 3 doi: 10.5376/pgt.2021.12.0003
Received: 01 Sep., 2021 Accepted: 06 Sep., 2021 Published: 09 Oct., 2021
Li Y.J., Qi S.J., Song X.Q., Zhang C.L., Ren M.X., and Yu X.D., 2021, Clonal growth characteristic of epiphytic orchid Dendrobium sinense, Plant Gene and Trait, 12(3): 1-8 (doi:10.5376/pgt.2021.12.0003)
Dendrobium sinense is a perennial epiphytic orchid, which belongs to the genus of Dendrobium and the family of Orchidaceae. Under natural conditions, D. sinense has a low seed setting rate and the environmental requirements for its seed germination a very stringent. Besides sexual propagation, asexual ways like clonal propagation is also found in D. sinense. In this study, five transects were set up in Bawang Ridge of Hainan Province to monitor the dynamic change of D. sinense population and study the influence of clonal propagation on population ecology of D. sinense. The main results are as follows: D. sinense belongs to a bulbous clonal plant, Spacer between pseudobulbs is only 0.35 cm around. Buds are tillered on the base of the one-year pseudobulb and deveiopment through the energy transfer between the ramets to continuation the genet life. Branching intensity enhanced as genet grow up. And different ramet from the genet have big difference in growth situation. The scale of genet cannot expansion infinitely, D. sinense can adjust expend speed according to the density of ramets. Clonal growth provides material and energy reserves to sexual reproduction and create more sites for flowering by increasing the growth axis.
Clonal plants are plants that spontaneously produce new individuals with the same genetic material as themselves under natural environmental conditions (Jackson et al., 1985). Clonal plants keep expanding-foraging behavior (Dong, 1996) and a broad spectrum of ecological countermeasures (Liu and Zhong, 1995), with higher utilization of water, light energy and carbon dioxide (Jiang and Dong, 2000). Therefore, cloned plants have strong survival competitiveness and adaptability to harsh environments, and play an important role in the process of ecological restoration and renewal (Chen et al., 1997). At present, the research objects of clone growth are mainly in the clone plants of herbaceous, semi-shrub, shrub and tree (Dong et al., 1999, 2000; Zhang et al., 2009). Epiphytes are very important in the tropical rain forest, with high plant diversity, but the origin and maintenance of diversity are not clear, so research on the clonal growth of epiphytes and its main influencing factors may be an important aspect to understand the formation and maintenance of epiphytes diversity.
Orchids are the most important group of epiphytes, with at least 25,000 species (Gravendeel et al., 2004), of which more than 70% are epiphytes. In addition, also includes the terrestrial orchids and the saprophytic orchids, two kinds of different way of life. There are different types of asexual reproduction in orchids for different lifestyles. Terrestrial orchids are dominated by tillering and rhizomes. The Cypripedium lentiginosum sprouts short rhizomes and new buds at the base of the plant before drying up (Liu et al., 2008). Tiller propagation is the main asexual propagation mode of Paphiopedium purpuratum, which produces tiller buds from the axils of the seedlings when they grow to 3-4 leaves (Liu et al., 2004). Saprophytic orchids are mostly cloned and grown by underground vegetative propagation stems (Zhou, 2005). Pseudobulbs of epiphytic orchid are vegetative storage organs (Ng and Hew, 2000), and also the main organs for clone growth.
D. sinense is endemic to Hainan and naturally distributed in the mountainous areas of central and western Hainan above 1,000 m (Song, 2005). It is used as a traditional medicine locally in Hainan (Jiang et al., 2011). The seed setting rate of D. sinense was affected by pollinators (Wu, 2013), and its seedlings had low germination rate, weak reproductive ability and slow growth under natural environment. D. sinense individuals must have 4 or more pseudobulbs before sexual reproduction, resulting in a gradual increase of resource investment in sexual reproduction (Qi et al., 2017). However, cloning and reproduction occurred before sexual reproduction, which was the main way to maintain the population of D. sinense (Wang et al., 2018). This study monitored the wild populations of D. sinense in Dongwu area of Bawang mountain, Hainan province from 2011 to 2016, and revealed the law of clonal growth of D. sinense, so as to answer the following scientific questions: (1) the type of clonal growth of D. sinense; (2) What are the growth rules of pseudobulbs and genets of D. sinense; (3) How to coordinate clonal growth and sexual reproduction of D. sinense.
1 Results and Analysis
1.1 Cloning and breeding methods and types of D. sinense
The pseudobulbs of D. sinense stand erect or curved upward in a rod-like shape. Like all kinds of pseudobulbs of Dendrobium, the adult plants can reach more than 15 cm, the stem is 3~4 mm thick, unbranched, and the longest one has 9 nodes with about 1.5~3 cm interval. The D. sinense grows two or more new buds to the base of the adult pseudobulbs with the help of the nutrition of the adult plants. The new buds absorb the nutrition of the old pseudobulbs to complete the energy accumulation and then sprout new shoots from the base to complete the clonal growth cycle (Figure 1A). Therefore, according to the classification of cloned organs, D. sinense belongs to the bulbous clonal plant of branch source clonal plants. The pseudobulbs of D. sinense were able to grow and complete the life history independently after being separated from the mother plant. The reason is that the pseudobulbs have special morphological structure and physiological function. The spacer was very short, the average distance was only (0.35±0.10) cm, and the branching angle varied widely. Because of the long life of the spacer and its clumps distribution, the clonal propagation of D. sinense is very intensive.
|
Figure 1 Clonal growth of pseudobulbs of D. sinense Note A: Budding; B: Two clone growth axes |
.png)
1.2 Branch intensity of D. sinense
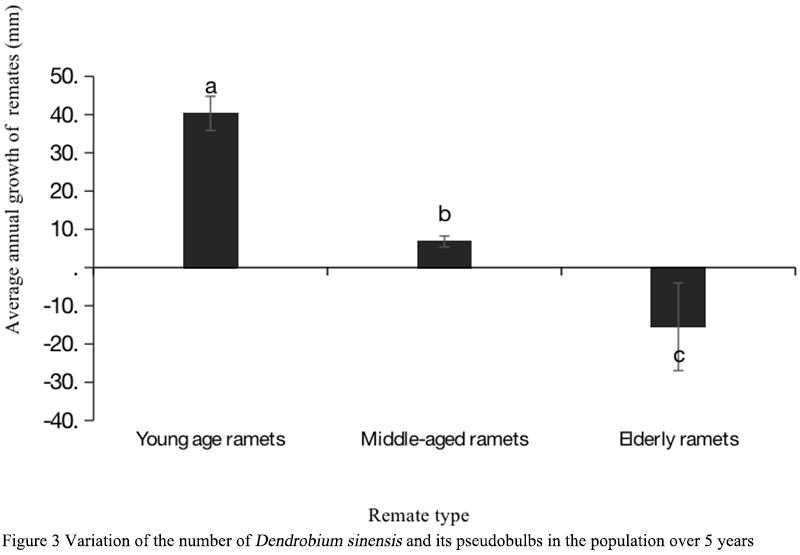
Down before and after each year in April, a part of the base of annual pseudobulbs shoots out 1~2 shoot buds and, in most cases, only one can expand into a new pseudobulb, but in rare cases there are also two to expand and grow into two new pseudobulbs, absorb the new pseudobulb through aging pseudobulb nutrition and germinate two or more growth axes in base, thus by a growth axis turning into two even more growth axes (Figure 1B). The number of axes increased with the increase of the age of D. sinense (Figure 2; Figure 3). Therefore, it can be inferred that the branch intensity of D. sinense is not constant, but increases with the increase of age class, so the new branch does not grow at a uniform rate.
|
Figure 2 Relationships of Age-class of genet with number of growth axes |
.png)
|
Figure 3 Variation of the number of Dendrobium sinensis and its pseudobulbs in the population over 5 years |
1.3 Expansion capacity of D. sinense
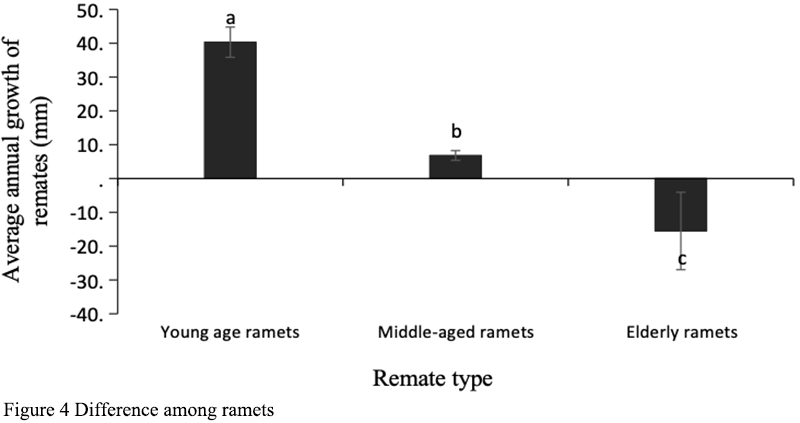
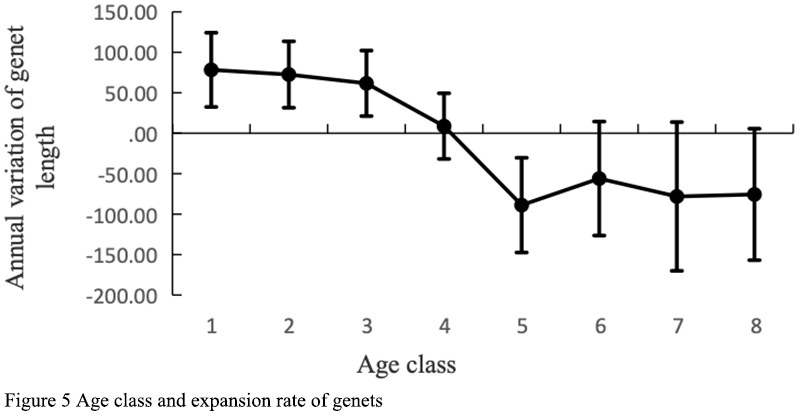
The expansion ability of clonal plant base strains is reflected in the number of plants produced in their life cycle and their occupation of horizontal space (Dong, 2011). D. sinense, as an intensive clonal plant, has limited horizontal expansion ability. As epiphytic plants on trees, a clonal cluster can occupy less than 0.05 m3 at most. The growth of different age remates in the same genet was different, the new and young pseudobulbs grew the fastest, while the mature pseudobulbs in the middle of the genet had limited growth, while the old pseudobulbs at the tail of the genet began to dry up and die (Figure 4). The growth status of the genet is not consistent with the life status of each remate, which is the comprehensive embodiment of each remate. We used the difference of the sum of all pseudobulb lengths of the same genet in the two years as the expansion rate of D. sinense. The size of the genets increased with the increase of the number of remates, but the rate of expansion was not constant. The early seedlings of D. sinense were not mature enough for sexual reproduction. With the growth of D. sinense, sexual reproduction did not occur until the individual reached a certain critical value, thus the expansion rate of D. sinense was slowed down. As the genets continued to grow, their sexual reproduction also increased due to multiple growth axes. At this time, the clone expansion rate of the genets slowed down significantly or even showed negative growth. At this time, the growth rate of the younger remates was not as fast as that of the older remates (Figure 5).
|
Figure 4 Difference among ramets |
|
Figure 5 Age class and expansion rate of genets |
1.4 Clonal growth and sexual reproduction
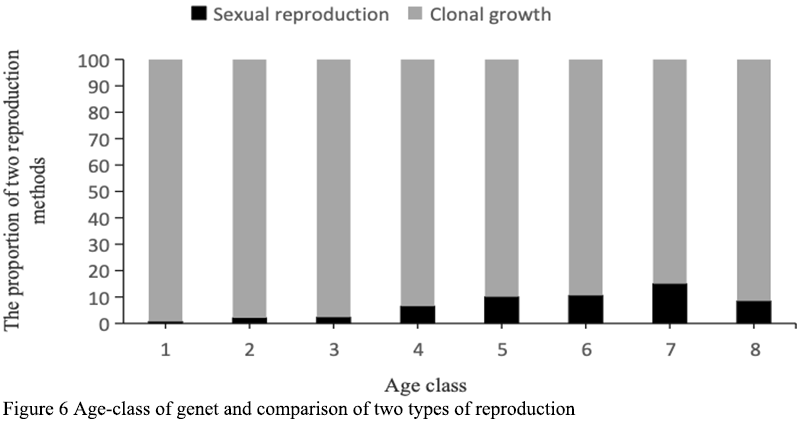
There are two breeding methods: sexual reproduction and clonal growth. According to field observation in the five years from 2011 to 2015, sexual reproduction could only be carried out when the individual size reached 4 pseudobulbs, and the resource allocation to sexual reproduction increased with the increase of age level (Figure 6). With the whole life cycle of the clonal growth, the seedling of D. sinense increased the number of remates and expanded the size of genets, and accumulated material and energy to prepare for sexual reproduction. In the late growth stage, D. sinense provided sites for sexual reproduction by increasing the branch strength and increasing the growth axis. Although clonal reproduction rarely increases the number of individuals in the population and only extends the life of the genets, the occurrence probability in the population can reach 100%, and far exceeds the resource investment of sexual reproduction. In this study, two propagation modes of D. sinense from 8 age classes were investigated. The proportion of sexual reproduction mode increased with the increase of age classes, and reached the highest proportion at 7 class (14%), and then began to decrease at 8 class (Figure 6).
|
Figure 6 Age-class of genet and comparison of two types of reproduction |
2 Discussion
D. sinense is a perennial plant. Its growth and reproduction are related to local climatic conditions. It adapts to seasonal drought with seasonal deciduous leaf, and relies on the fuzz on the stem to resist the cold climate. The stem of D. sinense is short and concentrated, which is conducive to improving drought tolerance and increasing their fitness in adversity (Hew and Ng, 1996). The extremely low seed setting rate of D. sinense was caused by the restriction of pollinators due to the synchronization of flowering period with the frequent typhoon period in Hainan Island (Zhou, 2013). Seed germination is a vulnerable stage in the life history of wild orchids, which in most cases requires clonal reproduction to maintain and expand population numbers (Zotz, 1998; Liu et al., 2010). The interconnection between the genet and the remates is the advantage of clonal reproduction. The material transfer between the spacers provides material guarantee for the survival of the seedlings, extends the life span of D. sinense and maintains the dynamic stability of the population. The clonal growth of D. sinense can effectively reduce the death of individuals and store nutrients, and plays an important role in the vegetative growth and sexual proliferation of D. sinense (Wu et al., 2013).
D. sinense is a typical epiphytic orchid plant. The ecological factors in the microenvironment of epiphytic plants have no obvious distribution rule and water and nutrients are very scarce, which indicates that the growth environment of D. sinense can more stimulate its plasticity phenotype (Dong, 2011). The growth types of D. sinense are changed according to the change of habitat and individual development stage, the angle changes between different remates were positively correlated with the branch strength the angle range increases, and the branch strength will increase too. In addition, the sites of sexual reproduction and feeding increase with the growth axis. The intensive cloning pattern of D. sinense results in more intraspecific and interspecific contact opportunities, thus increasing the degree of competition. the expansion speed of D. sinense is related with intraspecific competition, and with the number of remates of a negative correlation relationship. Plants use the integral effectiveness of cloning in order to reduce the competition of their remates, and adjust the life activities of growing point through the law of the density restriction mechanism, which can effectively adjust the density of remates. Some studies have found that clonal plants take measures to control the activity of growing points to avoid self-thinning (Pitelka et al., 1997). On the one hand, this mechanism ensures a reasonable population density, and on the other hand, it can improve the utilization rate of space and reduce the competition between remates.
Most clonal plants are capable of both clonal and sexual reproduction. The clonal growth of D. sinense provides material reserve and reproductive site for sexual reproduction, and sexual reproduction distributes excess material energy to regulate plant density. In addition, the distribution ratio of the two breeding methods was allocated according to the efficiency of sexual reproduction. In the wild environment, the two methods of asexual reproduction Cypripedium japonicum Thunb and Calanthe tsoongiana were adopted to ensure population reproduction and solve the problem of low flowering and seed setting rate (Liu et al., 2013). Clonal growth can maintain the reproductive capacity of the population and achieve the effect of sexual reproduction. Therefore, D.sinense still retains the mode of sexual reproduction, and develops by sexual reproduction when the habitat is suitable, but the proportion of sexual reproduction would be very small when the habitat is not suitable.
Although the clonal growth of many orchidaceae plants, including D. sinense, does not increase the population number, it plays an important role in extending the individual life span and maintaining the stability of the population. This study only revealed the types of clonal growth of D. sinense, the growth rate of remates and genets, and the relationship between the two propagation modes. There are still many scientific problems to be solved, such as the peak of flowering and seed setting rate of D. sinense with age level 7, which involves the tradeoff between reproduction and growth of D. sinense. In addition, D. sinense has germination differences among different epiphytic hosts (Yang et al., 2017), and whether there are differences in the growth of its clones on different epiphytic hosts or at different altitudes was the next research direction. In conclusion, this study provides a theoretical basis for the clonal growth of epiphytic orchids and lays a foundation for further discussion on the clonal laws of epiphytic orchids.
3 Materials and Methods
3.1 Overview of the research area
Located in the mountainous area in the southwest of Hainan Island (18.87°~19.20°N, 108.88°~109.33°E), Hainan Bawang National Nature Reserve is a nature reserve of forest ecosystem type. It is mainly mountainous with an altitude of 100~1654 m. The study area has a tropical monsoon climate with distinct wet and dry seasons (May ~ October) and dry seasons (November~April of the following year). The average annual temperature is 24.2℃ and the average annual precipitation is 2500 mm.
3.2 Data statistics
In This study, five 5 m×50 m sample strips were selected in September 2011 in The Dongwu area (Bawangling National Nature Reserve, Hainan), and the D. sinense samples in the area were marked. The length of the longest pseudobulbs in each individual was measured in September, the number of pseudobulbs was counted, the number of pseudobulbs with leaves and the number of flowering were recorded respectively, and then the number of fruit pods was counted at the end of October. Since 2015, some D. sinense plants in 5 sample strips (50 plants were randomly selected for each sample strip) were selected and the length of pseudobulbs was recorded in detail so as to calculate the length of clonal propagation in one year.
3.3 Age index and class classification
The statistical method of this study was further improved on the basis of the research of our group (Wu, 2013), and the age class index (age class index) was constructed based on the reproductive characteristics and growth mode, where the age class index Size index = the total number of pseudobulbs + the longest pseudobulb length /10. For the convenience of statistics, D. sinense was divided into 8 classes according to the age index (Table 1), and the different remates in the same genet were divided into three categories: young, middle age and old age (Table 2).
|
Table 1 Different age class of individuals for D. sinense |
.png)
|
Table 2 Ramet division of D. sinense |
.png)
3.4 Comparison of the two breeding methods
According to the method of Zheng (2004), the ratio of dry matter weight of sexual reproductive organs (Flowers, fruits) and vegetative reproductive organs (new remates) was used as the ratio of compounds consumed in sexual reproduction and clonal reproduction in one year, and the distribution law of the two reproductive modes in different age stages. In this study, 8 dendrobium individuals of different sizes in Dongwu area were randomly selected and the length of all green branches were measured. The annual and perennial pseudobulbs were differentiated, and their fresh weight were weighed. Then they were dried at a constant temperature of 85℃ and weighed with a one over ten-thousandth balance until the constant weight is reached. In addition, 5 flowers in full bloom and 5 fruits in mature stage were selected respectively and dried to constant weight under the same conditions above. The specific gravity of the two breeding methods of D. sinense was calculated according to the following formula:
Reproductive ratio = sexual reproductive organ biomass / vegetative reproductive organ biomass
Sexual organ biomass = annual number of flowering * single flower biomass + annual number of fruiting * single fruit biomass
Vegetative reproductive organ biomass = annual growth of new remates length *(annual remates biomass / annual plant length)
3.5 Data analysis
All experimental data in this study were analyzed and processed by Excel 2013 and SPSS 21.0.
Authors’ contributions
Li Yijia is the experimental designer and executor of this research. Li Yijia, Qi Shanjiang and Zhang Cuili completed data analysis and wrote the first draft of the paper. Yu Xudong and Ren Mingxun participated in experimental design and analysis of experimental results; Song Xiqiang was the initiator and leader of the project, directing experimental design, data analysis and paper writing and modification. All authors read and approved the final manuscript.
Acknowledgments
This study was funded by Hainan Natural Science Foundation Innovation Research Team project (2018CXTD331) and 2020 Young and Middle-aged Teacher Educational Research Project in Fujian [Technology] (JAT201268).
Chen S., Li Z.Z., and Wang G., 1997, Advances in researches of growth form of the clonal plant, Shengtaixue Zazhi (Chinese Journal of Ecology), 16(4): 59-63
Dong M., 1996, Clonal growth in plants in relations to resource heterogeneity: foraging behavior, Zhiwu Xuebao (Acta Botanica Sinica), 38(10): 828-835
Dong M., Alateng B., Xing X.R., and Wang Q.B., 1999, Genet features and ramet population features in the rhizomatous grass species Psammochloa villosa, Zhiwu Shengtai Xuebao (Acta Phytoecologica Sinica), 23(4): 302-310
Dong M., ed, 2011, Clonal plant ecology, Science Press, Beijing, China, pp.66
Dong M., Zhang S.M., and Chen Y.F., 2000, Clonal plasticity in response to nutrient availability in the stoloniferous herb, Duchesnea indica, Zhiwu Xuebao (Acta Botanica Sinica), 42(5): 518-522
Gravendeel B., Smithson A., Slik F.J.W., and Schuiteman A., 2004, Epiphytism and pollinator specialization: drivers for orchid diversity?, Philos. Trans. R. Soc. Lond., B, Biol. Sci., 359(1450): 1523-1535
https://doi.org/10.1098/rstb.2004.1529
PMid:15519970 PMCid:PMC1693444
Hew C.S., and Ng C.K.Y., 1996, Changes in mineral and carbohydrate content in pseudobulbs of the C3 epiphytic orchid hybrid Oncidium Goldianaat at different growth stages, Lindleyana, 11: 125-134
Jackson B.C., Buss L.W., and Cook R.E., 1985, Population biology evolution of clonal organisms, In: Silander J.A.J., ed., Microevolution in clonal plants, Yale University Press, New Haven, USA, pp.105-152
Jiang G.M., and Dong M., A comparative study on photosynthesis and water use efficiency between clonal and non-clonal plant species along the northeast China transect (NECT), Zhiwu Xuebao (Acta Botanica Sinica), 42(18): 855-863
Jiang H.Y., Yang F.S., and Song X.Q, 2011, Preliminary study on artifical sowing of Dendrobium sinense in natural habitat in Bawangling natural reserve, Hainan Island, Redai Linye (Tropical Forestry), 39(2): 40-43
Liu F., Li J.Q., Wang C.X., Lian J.J., and Tian M., 2013, Floral characteristics and breeding systems of an endangered species Cypripedium japonicum, Linye Kexue (Scientia Silvae Sinicae), 49(1): 53-60
Liu Q., and Zhong Z.C., 1995, Advances in ecological research of clonal plant population and some related concepts, Shengtaixue Zazhi (Chinese Journal of Ecology), 14(3): 40-45
Liu Q., Yin S.H., and Lan Q.Y., 2010, Research advances in population dynamics of Orchidaceae, Yingyong Shengtai Xuebao (Chinese Journal of Applied Ecology), 21(11): 2980-2985
Liu Z.J., Chen L.J., Rao W.H., Li L.Q., and Zhang Y.T., 2008, Correlation between numeric dynamics and reproductive behaviour in Cypripedium lentiginosum, Shengtai Xuebao (Acta Ecologica Sinica), 28(1): 111-121
https://doi.org/10.1016/S1872-2032(08)60021-9
Liu Z.J., Zhang J.Y., Ru Z.Z., Lie S.P., and Chen L.J., 2004, Conservation biology of Paphiopedilum purpuratum (Orchidaceae), Shengwu Duoyangxing (Biodiversity Science), 12(5): 509-516
Ng C.K.Y., and Hew C.S., 2000, Orchid pseudobulbs-‘false’ bulbs with a genuine importance in orchid growth and survival!, Sci. Hortic., 83(3-4): 165-172
https://doi.org/10.1016/S0304-4238(99)00084-9
Pitelka L.F., 1977, Energy allocation in annual and perennial lupines (lupinus: Leguminosae), Ecology, 58(5): 1055-1065
https://doi.org/10.2307/1936925
Qi S.J., Meng Q.W., Song X.Q., Zhang Z., Hu X.Y., and Yu X.D., 2017, Reproductive allocation of Dendrobium sinense, an endemic orchid species to Hainan Island, Redai Shengwu Xuebao (Journal of Tropical Biology), 8(3): 313-317
Song X.Q., 2005, Studies on the investigation of wild Dendrobium germplasm resources in Hainan Island with special reference to conservation biology of D. sinense, Dissertation for Ph.D., Beijing Forestry University, Supervisor: Zhang Q.X., pp.1-10
Wang T.X., Qi S.J., Song X.Q., Meng Q.W., and Yu X.D., 2018, Correlation analysis between population dynamics and reproduction strategies in Dendrobium sinense (Orchidaceae), an endemic orchid in Hainan Island, Redai Shengwu Xuebao (Journal of Tropical Biology), 9(2): 189-197
Wu H.Z., 2013, Population ecology of Dendrobium sinense (Orchnidaceae), an endemic species to Hainan Island, Thesis for M.S., Hainan uinversity, Supervisor: Zhu G.Pand Song X.Q., pp.13-45
Wu H.Z., Song X.Q., Yang F.S., and Zhu G.P., 2013, Pseudobulbs on the survival and mating system of Dendrobium sinense (Orchidaceae) in Hainan Island, Redai Zuowu Xuebao (Chinese Journal of Tropical Crops), 34(4): 625-629
Yang F.S., Sun A.H., Zhu J., Jason D., Song X.Q., and Liu H., 2017, Impacts of host trees and sowing conditions on germination success and a simple ex situ approach to generate symbiotic seedlings of a rare epiphytic orchid endemic to Hainan Island, China, Bot. Rev., Doi: 10.1007/s12229-017-9178-1
https://doi.org/10.1007/s12229-017-9178-1
Zhang D.Y., Wang J.C., and Shi X., 2009, Poulation quantitative dynamics of the rhizomatous woody clonal plant Eremospartion songoricum in China’s curbantggut desert, Zhiwu Shengtai Xuebao (Acta Phytoecologica Sinica), 33(5): 893-900
Zheng W., 2004, Reproductive allocation of two clonal plants: the influence of grazing, fertilization and light, Thesis for M.S., Xinjiang Agricultural University, Supervisor: Zhu Z.H., and Zhu J.Z., pp.29-42
Zhou Y., 2005, Gastrodia biological characteristics, Thesis for M.S., Northwest A&F University, Supervisor: Mu X.Q., pp.11-15
Zotz G., 1998, Demography of the epiphytic orchid, Dimerandra emarginata, J. Trop. Ecol., 14(6): 725-741



. PDF(1393KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Yijia Li
. Shanjiang Qi
. Xiqiang Song
. Cuili Zhang
. Mingxun Ren
. Xudong Yu
Related articles
. Dendrobium sinense
. Population ecology
. Clonal growth
. Sexual reproduction
Tools
. Email to a friend
. Post a comment