Hydrogen Peroxide Priming Stimulates Drought Tolerance in Mustard (Brassica juncea L.) Seedlings 
Mohammad Anwar Hossain
1,2 
,
Masayuki Fujita
2 
1.Laboratory of Plant Stress Responses, Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, Miki-cho, Kita-gun, Kagawa-761-0795,Japan
2.Department of Genetics and Plant Breeding, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh
Author
Correspondence author
Plant Gene and Trait, 2013, Vol. 4, No. 20 doi: 10.5376/pgt.2013.04.0020
Received: 03 Jul., 2013 Accepted: 17 Aug., 2013 Published: 05 Oct., 2013
© 2013 BioPublisher Publishing Platform
This is an open access article published under the terms of the
Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Preferred citation for this article:
Hossain and Fujita, 2013, Hydrogen Peroxide Priming Stimulates Drought Tolerance in Mustard (Brassica juncea L.) Seedlings, Plant Gene and Trait, Vol.4, No.20 109-123 (doi: 10.5376/pgt.2013.04.0020)
Abstract
Hydrogen peroxide, a central modulator of stress signal transduction pathways, activates multiple defense responses that reinforce resistance to various abiotic and biotic stresses in plants. The present study examines the potential biochemical mechanisms of hydrogen peroxide pre-treatment-induced drought tolerance in mustard (Brassica juncea L.) seedlings by analyzing numerous vital components of methylglyoxal and reactive oxygen species detoxification systems. Eight-day-old seedlings were pre-treated with low concentration (50 µM) of hydrogen peroxide for 24 h prior to the imposition of drought stress (20% PEG-6000) for 48 h. Hydrogen peroxide pre-treatment enhanced the membrane stability of leaf tissues under drought stress, as revealed from greatly reduced malondealdehyde content. The level of endogenous hydrogen peroxide contents in exogenous hydrogen peroxide pre-treated drought stressed-seedlings was markedly lower than those of the seedlings subjected to drought stress without hydrogen peroxide pre-treatment. A declination in the activities of ascorbate peroxidase, catalase and glyoxalase II were observed in response to drought stress whereas dehydroascorbate reductase, glutathione peroxidase and glyoxalase I activities significantly increased. The content of ascorbate, reduced glutathione and oxidized glutathione increased significantly whereas glutathione/glutathione disulphide ratio decreased in drought-stressed seedlings. Surprisingly, hydrogen peroxide pre-treated drought-stressed seedlings maintained a significantly higher ascorbate peroxidase, glutathione reductase, catalase, glutathione S-trasnferase, and glyoxalase II activities and glutathione/glutathione disulphide ratio when compared with the seedlings subjected to drought stress without hydrogen peroxide pre-treatment. Our results indicated that hydrogen peroxide primed a defense response in the seedlings that could trigger the activation of both ROS and MG detoxification pathways and enabled the seedlings tolerance to drought-induced oxidative damage.
Keywords
Drought stress; Reactive oxygen species; Methylglyoxal; Hydrogen peroxide; Mustard
In their natural habitat plants regularly experience multiple abiotic and biotic stresses from which they cannot escape and for which they have evolved intricate mechanisms to detect environmental changes, allowing optimal responses to adverse conditions (Krasensky and Jonak, 2012). Drought or water stress is a major abiotic agent that seriously reduces crop productivity and crop expansion worldwide (Yang et al., 2010). Predicted changes in climatic variability and rainfall pattern are expected to make crop improvement even more crucial for food production (Lamb, 2012). Therefore, the development of drought tolerant crop varieties has become an urgent concern for many crop-breeding programs to ensure global food security. Although improved adaptation to abiotic stress has long been a pursuit of breeders, it has been difficult to achieve due to the multigenic origin of the adaptive responses (Lopes et al., 2011). One of the biggest challenges to modern sustainable agricultural development is to obtain new knowledge that could allow breeding and engineering of plants with new and desired agronomical traits (Le et al., 2007; Duque et al., 2013). Therefore, the importance of understanding the molecular and biochemical basis of drought stress responses and tolerance is driven by both an interest in basic knowledge and the prospect that such knowledge might provide new strategies for drought stress tolerance in plants for more sustainable agricultural production.
Water stress, salinity stress, extreme temperatures and oxidative stress are often interconnected, and may induce similar cellular damage (Wang et al., 2003; Singh et al., 2011). Upon exposure to water stress, plants dramatically increase the production of reactive oxygen species (ROS) such as singlet oxygen (1O2), superoxide (O2•−), hydrogen peroxide (H2O2), and the hydroxyl radical (•OH) in different subcellular organelles (Cruz de Carvalho, 2008). Restricted entries of CO2 in the leaves during drought stress limit the CO2 fixation and accelerate the photorespiratory pathway and finally lead to excessive H2O2 in the peroxisome (Cruz de Carvalho, 2008). It has been estimated that under drought stress more than 70% of total H2O2 accumulation is due to photorespiration (Noctor et al., 2002). Although ROS mainly H2O2 can act as signals to help plants adapt to stress responses (Miller et al., 2008, 2010; Gill and Tuteja, 2010; Hossain and Fujita, 2012; Petrov and Van Breusegem, 2012), excess ROS cause oxidative damages to plant macromolecules (Gill and Tuteja, 2010; Jaspers and Kangasjarvi, 2010; Hossain et al., 2011a). As a result metabolic alteration, inhibition of photosynthesis, and breakdown of cellular organization contribute to growth retardation, reduced fertility, premature senescence even death of plants (Saito et al., 2010; Krasensky and Jonak, 2010; Hossain et al., 2011a). Therefore, the level of ROS should be judiciously regulated in plants through the coordinate of ROS production system and ROS scavenging system to manage oxidative damage and simultaneously regulate signalling events (Foyer and Noctor, 2005; Rouhier et al., 2008), but the fundamental mechanisms are still largely unknown.
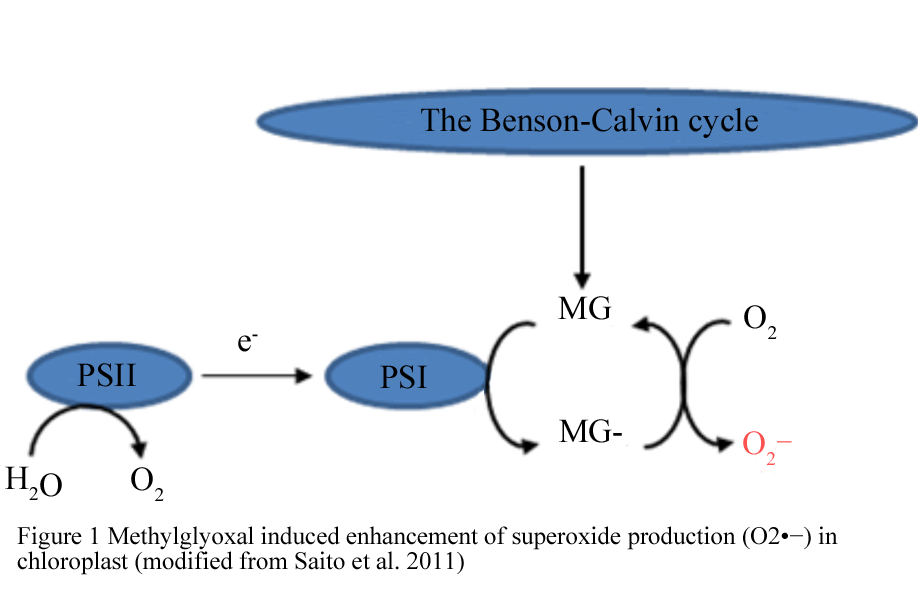
Methylglyoxal (MG) and abiotic stresses are inextricably intertwined, the over-accumulation and higher rate of MG production in plants possibly being a general stress response (Hossain et al., 2009; Banu et al., 2010; Upadhyaya et al., 2011). MG is a typical α-oxoaldehyde, which forms as a by-product of several metabolic pathways, e.g. glycolysis, lipid peroxidation and oxidative degradation of glucose and glycated proteins. It is toxic to plant cells, causing inhibition of cell proliferation, degradation of proteins and inactivation of antioxidant defense systems and as a consequence disrupts cellular functions (Matins et al., 2001; Hoque et al., 2010, 2012a). MG accumulates in plants under normal plant physiological processes like photosynthesis and MG vividly elevated under various stress conditions (Yadav et al. 2005a; Hossain et al., 2009). Recently Saito et al. (2011) reported that MG accumulated in chloroplasts during the day from triose phoshphates, needs to be controlled by detoxification mechanisms, otherwise it will catalyse the photoreduction of O2 to O2•− at photosystem I (Figure 1) and the increase in O2•−production during photosynthesis further aggravate the oxidative damage to plant cells. MG not only directly inhibits physiological functioning but it also inhibits it via the changes in ABA synthesis in Arabidopsis (Hoque et al., 2012b, 2012c). Therefore, in order to survive under stressful conditions plants must up-regulate MG detoxification process to avoid cellular damage and also to keep steady state pace in different plant physiological processes.
|
.png)
Figure 1 Methylglyoxal induced enhancement of superoxide production (O2•−) in chloroplast (modified from Saito et al., 2011)
|
To avoid the excessive accumulation of ROS and to protect from oxidative damage, plants are armed with sophisticated antioxidant defense systems: both enzymatic antioxidant (multiple superoxide dismutases (SOD), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), glutathione reductase (GR), catalase (CAT), glutathione peroxidase (GPX), glutathioneS-transferase (GST), peroxidase (POX), guaiacol peroxidase (GPOX) and non-enzymatic antioxidant (asocrbare (AsA), glutathione (GSH), tocopherol, carotenoids, flavonoids, and proline) (Figure 2; Gill and Tuteja, 2010; Hossain et al., 2011a; Sharma et al., 2012) in different sub-cellular organelles. The delicate balance between ROS production and scavenging that allows this duality in function to exist in plants is thought to be orchestrated by a large network of genes that tightly regulates ROS production and scavenging (Apel and Hirt, 2004;
Mittler et al., 2004; Miller et al., 2008; Petrov and Van Breusegem, 2012).