Cloning and Analysis of Gene Fragments Encoding C4H and CAD from Miscanthus sinensis 
 Author
Author  Correspondence author
Correspondence author
Plant Gene and Trait, 2013, Vol. 4, No. 6 doi: 10.5376/pgt.2013.04.0006
Received: 17 Jul., 2013 Accepted: 19 Jul., 2013 Published: 23 Jul., 2013
Fan and Li, 2013, Cloning and Analysis of Gene Fragments Encoding C4H and CAD from Miscanthus sinensis, Plant Gene and Trait, Vol.4, No.6 30-36 (doi: 10.5376/pgt.2013.04.0006)
Based on cDNA sequences encoding Cinnamate-4-hydroxylase (C4H) and Cinnamoyl alcohol dehydrogenase (CAD) isolated from several monocots reported in GenBank, two pairs of PCR primers were designed with Primer Premier 5.0 software. The total RNA was extracted from Miscanthus sinensis according to CTAB-LiCl method and then cDNA was synthesized by reverse transcription. PCR products, which were named as MsC4H and MsCAD, were successfully obtained by reverse transcription polymerase chain reaction (RT-PCR) and then cloned into vector pMD18-T. The positive clones identified by plasmid PCR were sequenced. The sequencing results revealed that MsC4H and MsCAD contain 305 and 269 base pairs, which encode 101 and 51 amino acids, respectively. BLAST analysis showed that the cloned nucleotide sequences and their corresponding amino acids are highly homologous to many C4H and CAD genes and their encoding proteins from different plant species. The sequences of MsC4H and MsCAD had been submitted to Genbank database, whose accession numbers are JQ598686 and JQ598683, respectively.
Background
At present, 79% of the global energy consumption is met by fossil energy sources (crude oil, natural gas and coal) (Soetaert and Vandamme, 2006), which are the main energy sources to support the global world economic development and social progress. Due to limited reserves, it is generally agreed upon that petroleum will run out within 50 years, natural gas within 65 years and coal in approximately 200 years at the present rate of energy consumption. The rapid increase in fossil energy consumption has resulted in the environmental and climate problems such as the greenhouse gas emissions, air pollution and acid rain (Wuebbles and Jain, 2001). Therefore, countries around the world are looking for the renewable energy sources to meet the growing energy crisis and biomass is one of the most potential renewable energy sources. An ideal biomass crop is one with a high energy output whilst requiring minimal inputs (Jones and Walsh, 2001). Miscanthus sinensis belongs to the genus of Miscanthus, subfamily of Panicoidea, family of Poaceae (Zhan et al., 2012), which has a higher energy yield per hectare than other bioenergy crops such as willow, poplar, oil and starch crops (Hastings et al., 2008). Miscanthus has an efficient C4 photosynthesis process, use of water and nitrogen (Liu et al., 2011). Due to its low mineral content, full combustion, zero net effect of carbon dioxide, harmless gas emissions, helpful for the greenhouse effect alleviation and the soil and water conservation, resistent to drought, low dependence on water and fertilizer, easy to harvest (Heaton et al., 2004; Yuan et al., 2008), Miscanthus is destined to become one the most useful species among the second generation energy crops. The European Union aims to displace 10% of fossil fuels in the transportation sector with biofuels in 2020 (Fulton, 2005, http://www.ifri.org/files/Agriculture/FultontalkIFRI29March.pdf).
Although lignocellulosic biomass is the most promising feedstock for bioethanol production, large scale commercial production plants are not in use yet in the US or EU because of the high production costs (Sticklen, 2008). Procession lignocellulose into bioethanol occurs in four main steps: pretreatment, hydrolysis, fermentation and product separation. Pretreatment with chemical (acid), biological or mechanical method is the first step in order to reduce the size of the lignocellulosic material, hence increasing the hydrolysis rate of cellulose and hemicellulose in the second step (Mosier et al., 2005). Hydrolysis is to degrade the macromolecular cellulose and hemicellulose into monosaccharides, including hexose, xylose, mannose, galactose, arabinose and other oligosaccharides. Monosaccharide will be fermented into ethanol under the action of the enzymes. The last step involves a distillation and high purity ethanol will be achieved (Arshadi and Sellstedt, 2008). At present, among the industry chain from lignocellulosic biomass into bio-ethanol, a major limitation is the recalcitrance of lignin, which decreased the saccharification efficiency. Because the hydrolysis enzymes are expensive, thereby greatly increasing the cost of production of bio-ethanol (Chen et al., 2002).
Lignin, one kind of phenylpropanoid derivatives deposited on the cell wall of vascular plants, is composed of polymer compound with high molecule weight up to hundreds to millions connected by three lignin monomers by ether bond or a carbon-carbon bond, which accounts for 15%~35% weights of plants (Zhong et al., 2000). Lignin can enhance the mechanical strength of plants, improve cell transport capacity (Whetten and Sederoff, 1995), and can resist against pathogenic microorganisms (Duthie and Crozier, 2000). Lignin biosynthesis of vascular plants is an important evolutionary adaptation characteristic to terrestrial environment. Biosynthesis of lignin monomer is a very complex physiological and biochemical process in plant, which is the result of phenylpropanoid pathway conducted by deamination of phenylalanine (or tyrosine) into cinnamic acid, through a series of hydroxylation, methylation and reduction reactions, which ultimately produce three kinds of main monomer, namely syringyl lignin (S-lignin), guaiacyl lignin (G-lignin), p-hydroxyphenyl lignin (H-lignin) (Fan et al., 2005).
Cinnamate-4-hydroxylase (C4H) and cinnamoyl alcohol dehydrogenase (CAD) are two key enzymes of lignin biosynthesis (Vanholme et al., 2010; Hamada et al., 2004). In this experiment, the total RNA was extracted from Miscanthus with CTAB-LiCl method. Based on the cDNA sequences of C4H and CAD isolated from several monocots reported in GenBank, two pairs of PCR primers were designed. Cloning and bioinformatic analysis of cDNA fragments of C4H and CAD genes were carried out, hence laying a foundation for the next full-length cloning of C4H and CAD genes with RACE (rapid amplification of cDNA ends) technique and gene expression analysis with qPCR (Real-time Quantitative PCR) technique in the future.
1 Results and analysis
1.1 Extraction of total RNA
After the total RNA was extracted from Miscanthus with CTAB-LiCl method and then was digested with DNase I, the purified RNA was detected with the agarose gel electrophoresis (Figure 1). It indicated that two bands of 28S and 18S rRNA were clear enough, and the brightness of the 28S rRNA band to 18S rRNA is about 1.5 times, which showed that the extracted RNA, with high purity and high integrity, was suitable for the next step for reverse transcription and RT-PCR amplification.
|
|
.png)
1.2 RT-PCR amplification of the target genes
With the synthesized cDNA first strand as template and the two pairs of speicific PCR primers, two targed PCR products (Figure 2), named as MsC4H and MsCAD, were successfully amplified, respectively.
|
|
.png)
1.3 Characterization of the positive clones of the target genes
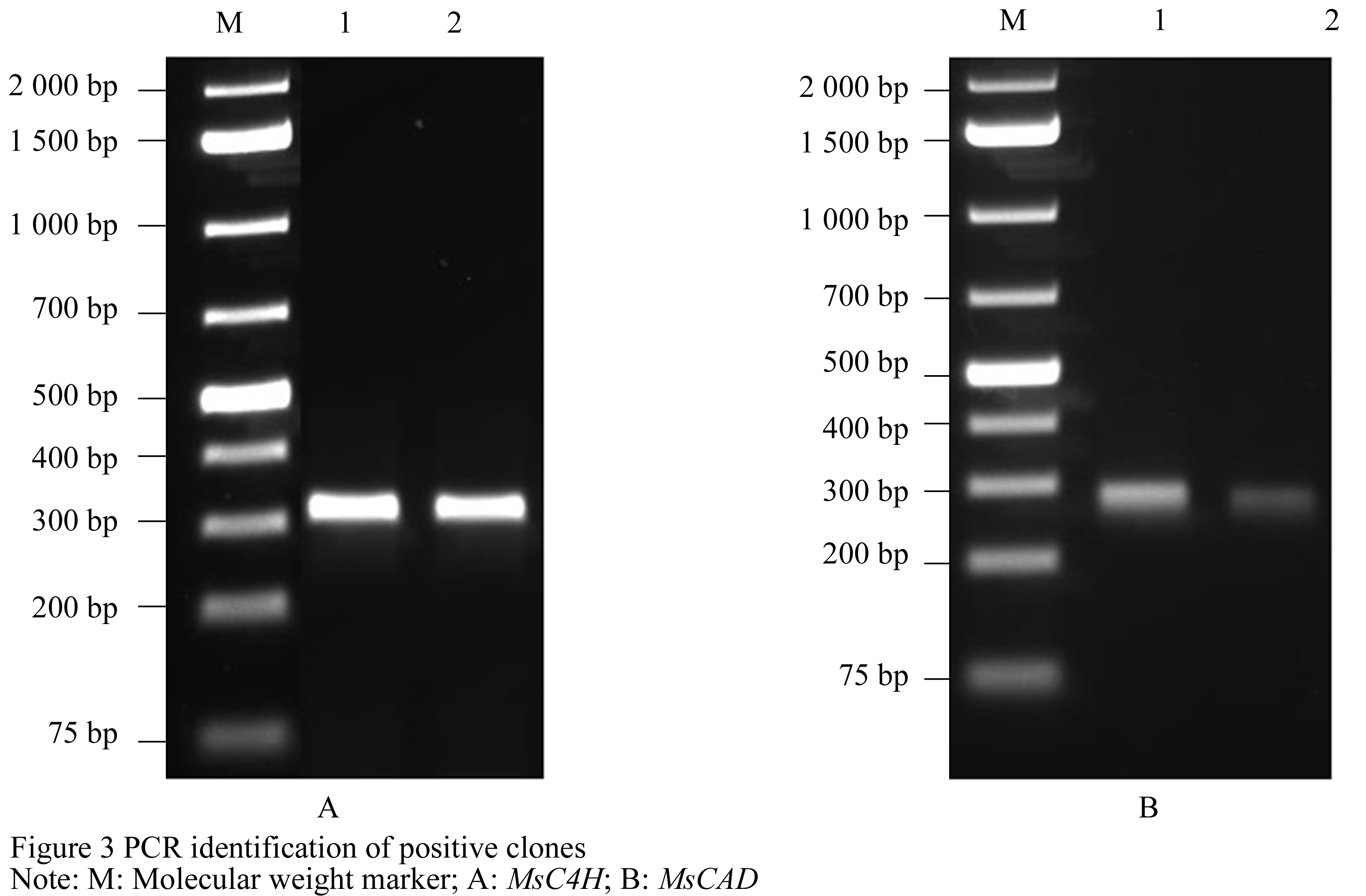
After the two targets PCR products were recovered from Agarose gel, they were ligated to pMD18-T and the ligated products were transformed into the competent cells of E. coli DH5α. The single clones of transformants were cultivated in LB liquid medium overnigh and then the plasmid werer isolated from E. coli for PCR identification. The PCR products were identical to the RT-PCR amplification (Figure 3), which indicated that they were positive recombinants.
|
|
1.4 Sequencing results and sequences analysis
Sequencing results demonstrated that the length of MsC4H and MsCAD fragment was 305 and 269 base pairs, which encoded 101 and 51 amino acids (Figure 4 and Figure 5), respectively. Blastn analysis revealed that the nucleotide sequences of MsC4H and MsCAD were highly homologous to many C4H and CAD cDNAs cloned from different species. The homology of MsC4H with C4H isolated from Sorghum bicolor (XM_002451999) and Zea mays (NM_001157893) was up to 98% and 94%. The homology of MsCAD with CAD cloned from Brachypodium distachyon was 80%. BlastP analysis revealed that the amino acid sequences encoding by MsC4H and MsCAD had high homology with C4H and CAD protein from many plants. The homology of C4H with C4H protein from Zea mays (NP_001151365) and Oryza sativa Japonica Group (NP_001046814, NP_001046819) were uo to 90%. The homology of CAD with CAD from Zea mays (NP_001131273, DAA35929) was more than 83%. Moreover, C4H protein contained CyPX (Cytochrome P450 monooxygenases) superfamily conserved domain, and CAD protein contained MDR (Medium-chain dehydrogenase/reductase) superfamily NAD(P) binding sites, which indicated the isolated genes in this experiment were true C4H and CAD genes. The sequences of MsC4H and MsCAD had been submitted to Genbank, whose accession numbers were JQ598686 and JQ598683.
|
|
|
|
2 Discussion
C4H was involved in the initiation reaction of biosynthesis of lignin monomer, which was the second enzyme in lignin metabolism pathway. Under the action of phenylalanine ammonia lyase (PAL), phenylalanine was deaminated to cinnamic acid, and then was converted to 4-coumaraic acid by C4H. After the methylation reaction and subsequent ligation reaction, three kinds of hydroxy cinnamic acid CoA esters were generated. The latest research demonstrated that saccharification efficency of the secondary wall of the antisense C4H transgenic tobacco was improved (Cook and Daudi, 2012), hence providing the new strategy and basis to improve the applications of the biomass crops in the industry.
CAD is the last enzyme in lignin monomer biosynthesis, involving the reduction reaction with Cinnamoyl-CoA reductase (CCR). CCR restores three kinds of hydroxy cinnamic acid CoA esters to form the corresponding cinnamic aldehyde, and then CAD deoxidized it to three kinds of cinnamyl alcohol. The enzyme activity of CAD of antisense CAD transgenic Medicago sativa L. was reduced dramatically. Although the lignin content remains unchanged, but the content of the S monomer decreased, resulting in S/G (syringyl/guaiacyl) ratio and S+G yield was significantly reduced. The digestibility of the genetically modified alfalfa by CAD gene has been improved. The enzyme activity of transgenic Populus tremula×Populus alba of antisense and sense CAD decreased 70% compared to the control, hence lowering the use of the chemicals to get rid of the lignin during the pulp and paper process and reducing the cost of pulp and paper.
Based on two pairs of the specific PCR primers, cDNA fragments of C4H and CAD were isolated from bioenergy plant, Miscanthus, therfby laying a foundation to clone the full-length cDNA sequences of C4H and CAD by RACE technique in the future. We also cloned the cDNA fragments encoding caffeoyl CoA 3-O-methyltransferase (CcoAOMT) (JQ598685) and peroxidase (POD) (JQ598684) from Miscanthus. We plan to carry out quantitative gene expression analysis of these four genes by qPCR technique, and analyze the correlation of mRNA transcription expression level of these genes with the saccharification efficiency, therefore providing the transformation target gene in order to orientedly molecularly improve the saccharification efficiency of the Miscanthus germplasm.
3 Materials and Methods
3.1 Materials
3.1.1 Plant material
The first internode of Miscanthus, plant material used in this experiment, was collected and frozen in liquid nitrogen, and then was taken to the laboratory and was immediately placed in -80℃ refrigerator.
3.1.2 Strain and vector
Escherichia coli strain DH5α was kept in our own laboratory. Cloning vector pMD18-T was bought from TaKaRa Biotechnology (Dalian) Co. LTD.
3.1.3 Reagent
Dnase I, M-MLV RTase cDNA Synthesis Kit, Taq DNA polymerase and Agarose Gel DNA Purification Kit were purchased from TaKaRa Biotechnology (Dalian) Co. LTD. Other biochemical reagents were all analytical grade reagents imported or domestically produced.
3.2 Methods
3.2.1 Total RNA extraction
The total RNA was isolated from Miscanthus according to the modified CTAB-LiCl method (Gasic et al., 2004). After the total RNA was digested by Dnase I to get rid of the residual trace DNA contamination, it was characterized by 1.2% agarose gel electrophoresis.
3.2.2 Primers design
Based on C4H and CAD sequences of several monocotes reported in Genbank, two pairs of PCR primers C4H-F (5'-CTCGCGCAGAGCTTCGACTA), C4H-R (5'-TCCCACTCGATGGACCAGAG) and CAD-F (5'-CGGAAGGAATCGACACAGAG), CAD-R (5'-GCACCAACCTGAGTTACAAC), were designed with Primer Premier 5.0 software. All the primers were synthesized by Sangon Biotech (Shanghai) Co. Ltd.
3.2.3 cDNA synthesis
cDNA was synthesized according to the protocol of M-MLV RTase cDNA Synthesis Kit, and then was placed in -20℃ refrigerator.
3.2.4 RT-PCR amplification
50 µL PCR reaction system was composed of 5 µL 10×PCR Buffer, 5 µL 2.0 mmol/L each dNTP, 1 µL cDNA, 1.5 µL 10 pmol/µL forward and reverse primer, respectively, 1 µL Taq DNA polymerase and 35 µL ddH2O. Thermocycling was performed at 95℃ for 5 min, then at 95℃ for 30 s, 54℃ for 30 s, 72℃ for 1 min for 35 cycles, 72℃ for 7 min. The PCR products were detected by 1% Agarose gel electrophoresis.
3.2.5 Characterization of the positive clones
The target PCR products were recovered with TaKaRaTM Agarose Gel DNA Purification Kit. Then the purified PCR products were ligated to pMD18-T and the ligated products were transformed into the competent cells of E. coli DH5α. The transformants were characterized by colony PCR and plasmid PCR. The positive recombinants were sequenced by Sangon Biotech (Shanghai) Co. Ltd.
3.2.6 Sequences analysis
The sequence homology analysis was carried out by online BLAST at NCBI (http://blast.ncbi.nlm.nih.gov/). The sequences were submitted to Genbank database with Sequin software.
Authors’contributions
FBY was the person who took charge of this project, including experiment design, data analysis, writing and modifying of the manuscript. LF conducted this experiment.
Acknowledgements
This study was financially supported by the program for young teachers in university and college of Henan province (2010GGJS-075). Authors appreciate for the advice and revising comment by the reviewers.
References
Arshadi M., Sellstedt A., 2008, Introduction to chemicals from biomass, Manhattan: John Wiley and Sons, pp. 79-87
Baucher M., Bernard-Vailhé M.A., Chabbert B., Besle J., Opsomer C., Montagu M.V., and Botterman J., 1999, Down-regulation of cinnamyl alcohol dehydrogenase in transgenic alfalfa (Medicago sativa L.) and the effect on lignin composition and digestibility, Plant Molecular Biology, 39(3): 437-447
http://dx.doi.org/10.1023/A:1006182925584
PMid:10092173
Baucher M., Chabbert B., Pilate G., Doorsselaere J.V., Tollier M.T., Petit-Conil M., Cornu D., Monties B., Montagu M.V., Inze D., Jouanin L., and Boerjan W., 1996, Red xylem and higher lignin extractability by down-regulating a cinnamyl alcohol dehydrogenase in poplar, Plant Physiology, 112(4): 1479-1490
PMCid:PMC158080
Chen L., Auh C., Chen F., Cheng X., Aljoe H., Dixon R.A., and Wang Z., 2002, Lignin deposition and associated changes in anatomy, enzyme activity, gene expression, and ruminal degradability in stems of tall fescue at different developmental stages, Journal of Agricultural and Food Chemistry, 50(20): 5558-5565
http://dx.doi.org/10.1021/jf020516x
PMid:12236679
Cook C.M., Daudi A., Millar D.J., Bindschedler L.V., Khan S., Bolwell G.P., and Devoto A., 2012, Transcriptional changes related to secondary wall formation in xylem of transgenic lines of tobacco altered for lignin or xylan content which show improved saccharification, Phytochemistry, 74: 79-89
http://dx.doi.org/10.1016/j.phytochem.2011.10.009
PMid:22119077 PMCid:PMC3657182
Duthie G., and Crozier A., 2000, Plant-derived phenolic antioxidants, Current Opinion in Clinical Nutrition and Metabolic Care, 3(6): 447-451
http://dx.doi.org/10.1097/00075197-200011000-00006
PMid:11085830
Fan B.Y., 2005, Regulation of lignin biosynthesis in transgenic plants by 4-coumarate: Coenzyme A ligase1 (4CL1) from Populus tomentosa and its enzymatic characterization, Dissertation for Ph.D., Beijing Forestry University, Supervisor: Jiang X.N., pp. 1-12
Gasic K., Hernandez A., and Korban S.S., 2004, RNA extraction from different apple tissues rich in polyphenols and polysaccharides for cDNA library construction, Plant Molecular Biology Reporter, 22(4): 437-438
http://dx.doi.org/10.1007/BF02772687
Hamada K., Nishida T., Yamauchi K., Fukushima K., Kondo R., and Tsutsumi Y., 2004, 4-coumarate: coenzyme A ligase in black locust (Robinia pseudoacacia) catalyses the conversion of sinapate to sinapoly-CoA, Journal of Plant Research, 117(4): 303-310
http://dx.doi.org/10.1007/s10265-004-0159-1
PMid:15235922
Hastings A., Clifton-Brown J., Wattenbach M., Stampfl P., Mitchell C.P., and Smith P., 2008, Potential of Miscanthus grasses to provide energy and hence reduce greenhouse gas emissions, Agronomy for Sustainable Development, 28: 465-472
http://dx.doi.org/10.1051/agro:2008030
Heaton E., Voigt T., and Long S. P., 2004, A quantitative review comparing the yields of two candidate C4 perennial biomass crops in relation to nitrogen, temperature and water, Biomass and bioenergy, 27(1): 21-30
http://dx.doi.org/10.1016/j.biombioe.2003.10.005
Jones M.B., and Walsh M., 2001, Miscanthus for energy and fibre, James & James (Science publishers), London, pp. 1-9
Liu M.X., Jiang J.X., and Yi Z.L., 2011, Application of cel l engineering in improving Miscanthus as a dedicated bioenergy crop, Caoye Xuebao (Acta Prataculturae Sinica), 20(4): 261-269
Mosier N., Wyman C., Dale B., Elander R., Lee Y.Y., Holtzapple M., and Ladisch M., 2005, Features of promising technologies for pretreatment of lignocellulosic biomass, Bioresource Technology, 96(6): 673-686
http://dx.doi.org/10.1016/j.biortech.2004.06.025
PMid:15588770
Soetaert W., and Vandamme E., 2006, The impact of industrial biotechnology, Biotechnology Journal, 1(7-8): 756-769
http://dx.doi.org/10.1002/biot.200600066
PMid:16897819
Sticklen M.B., 2008, Plant genetic engineering for biofuel production: Towards affordable cellulosic ethanol, Nature Reviews Genetics, 9(6): 433-443
http://dx.doi.org/10.1038/nrg2336
PMid:18487988
Vanholme R., Demedts B., Morreel K., Ralph J., and Boerjan W., 2010, Lignin biosynthesis and structure, Plant Physiology, 153(3): 895-905
http://dx.doi.org/10.1104/pp.110.155119
PMid:20472751 PMCid:PMC2899938
Whetten R., and Sederoff R., 1995, Lignin biosynthesis, The Plant Cell, 7: 1001-1003
http://dx.doi.org/10.2307/3870053
http://dx.doi.org/10.1105/tpc.7.7.1001
PMid:12242395 PMCid:PMC160901
Wuebbles D.J., and Jain A.K., 2001, Concerns about climate change and the role of fossil fuel use, Fuel Processing Technology, 71(1-3): 99-119
http://dx.doi.org/10.1016/S0378-3820(01)00139-4
Yuan J.S., Tiller K.H., Ai-Ahmad H., Stewart N.S., and Stewart C.N., 2008, Plants to power: Bioenergy to fuel the future, Trends in plant science, 13(8): 421-429
http://dx.doi.org/10.1016/j.tplants.2008.06.001
PMid:18632303
Zhan W.J., Ren J.X., Jin S.H., Huang Y.J., Pan Y.H., and Zheng B.S., 2012, Research progress on agronomic characteristics of Miscanthus, Zhejiang Nonglin Daxue Xuebao (Journal of Zhejiang A&F University), 29(1): 119-124
Zhong R., Morrison W.H. III, Himmelsbach D.S., Poole F.L. II, and Ye Z.H., 2000, Essential role of Caffeoyl coenzyme A O-methyltransferase in lignin biosynthesis in woody poplar plants, Plant Physiology, 124(2): 563-578
http://dx.doi.org/10.1104/pp.124.2.563
PMid:11027707 PMCid:PMC59163



. PDF(276KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Bingyou Fan
. Fang Li
Related articles
. Miscanthus sinensis
. C4H
. CAD , Cloning
. Sequence analysis
Tools
. Email to a friend
. Post a comment