Histological and Biochemical Changes in Aegle marmelos Corr. before and after Acclimatization 
2 Central Institute for Sub-tropical Horticulture, Rehmankhera, Lucknow-227107, India
 Author
Author  Correspondence author
Correspondence author
Tree Genetics and Molecular Breeding, 2013, Vol. 3, No. 3 doi: 10.5376/tgmb.2013.03.0003
Received: 18 Mar., 2013 Accepted: 31 Mar., 2013 Published: 27 Apr., 2013
Pati et al., 2013, Histological and biochemical changes in Aegle marmelos Corr. before and after acclimatization, Vol.3, No.3 12-18 (doi: 10.5376/tgmb. 2013.03.0003)
Maximum mortality of micropropagated plants occur during acclimatization phase because plantlets undergo rapid and extreme changes in physiological functioning, histological and biochemical changes. In order to investigate the actual reason of this limitation, test samples were collected at different stages of micropropagation of Aegle marmelos Corr. (in vitro stage , acclimation stage, field established plants). The biochemical result showed that micropropagated plantlets produced significantly low total chlorophyll (0.042 mg/g fresh weight), reducing sugar (3.227%), NR activity (1.353 NO2/h/g fresh weight) and but higher protein (0.048 µg/g) during in vitro phase. The in vitro raised plants showed abnormal histological features like altered leaf mesophyll, absence of thick cuticle, sunken stomata, poorly developed stem and root histology. These results revealed that the photoautrophic mode of nutrition during in vitro phase increased the survival rate during acclimatization compared to photoheterotrophic mode of nutrition. This suggests that photoautotrophism phenemoneon has substantial influence on the physiology and development of in vitro regenerated Aegle marmelos Corr. plantlets.
Introduction
Aegle marmelos Corr., belongs to family Rutaceae, is more prized for its pharmacological virtues than its edible quality. Because of pharmacological importance, it’s become potential candidate for developing transgenics to enhance its medicinal properties by researchers community (Pati and Muthukumar, 2013). But, commercial orcharding of Aegle marmelos Corr. despite its potential medicinal value, has not been taken up on large scale due to shortage of genuine planting materials for initial establishment of the orchard. Conventionally Aegle marmelos Corr., is propagated through grafting which is season dependent with lower multiplication rate. Micropropagation technique has been developed in Aegle mormelos Corr. from mature tree through enhanced axillary branching for mass multiplication (Pati et al., 2008; Pati et al., 2008a; Pati et al., 2008b; Raj and Basavaraju, 2012). However, high mortality of micropropagated plants during acclimatization and subsequently poor field establishment remains a major bottleneck in commercealizing the micropropagation protocol to industry. During in vitro condition plants grow in an atmosphere with high relative humidity, low light intensity, reduce gaseous exchange and low transpiration rates.
Plantlets often desiccate or wilt rapidly and finally die due to anatomical disturbances that affect the physiological process which includes, low photosynthetic rate, low transpiration rate, low uptake of water and minerals and high water loss during acclimatization. These anatomical and physiological disorders eventually affect the morphology of plantlets which lead to poor growth and low survival percentage of micropropagated plantlets (Kumar and Rao, 2012). Brainerd and Fuchigami (1982) reported that the characteristic inability of raised stomata to close upon removal could be corrected by acclimatization. Maren et al. (1970) observed that the reversion of stomata to functional state was achieved after removal from culture. Generally, in vitro conditions which promote rapid growth shoot proliferation and plantlet developed, resulted certain abnormal characteristics such as altered leaf morphology and mesophyll structure, poor photosynthesis, sunken and malfunctioning stomata (Ziv, et al., 1986; Hazarika, 1996). In order to devise an efficient acclimatization procedure, it was imperative to investigate some of the crucial biochemical (total chlorophyll, reducing sugar, NR activity and protein) and histological changes during various micropropagation stages, which contribute towards growth and development of plants.
1 Result and Analysis
1.1 Biochemical analysis of regenerated plants
The results revealed that a significant variation in biochemical parameters which were observed among the various culture stages. The chlorophyll (a, b and total) synthesis was recorded lower (0.022, 0.020 and 0.042 mg/g) during in vitro phase, while higher chlorophyll levels (0.093 mg/g) was recorded during acclimatization. The in vitro plants showed lower level (3.22 mg/g) of reducing sugars than ex vitro grown plants. Highest NR activity (1.35×10-2 NO2/h/g fresh weight) was observed during in vitro phase, which decreased significantly during acclimatization. The protein levels remain low during acclimatization (0.048 µg/g) as compared to control plant (Table 1).
| Table 1 Biochemical analysis of nodal stem segment-regenerated plants of Aegle marmelos. Different letters within column indicate statistically different values according to Tukey’ test (p=0.05) |
1.2 Histological study of leaf
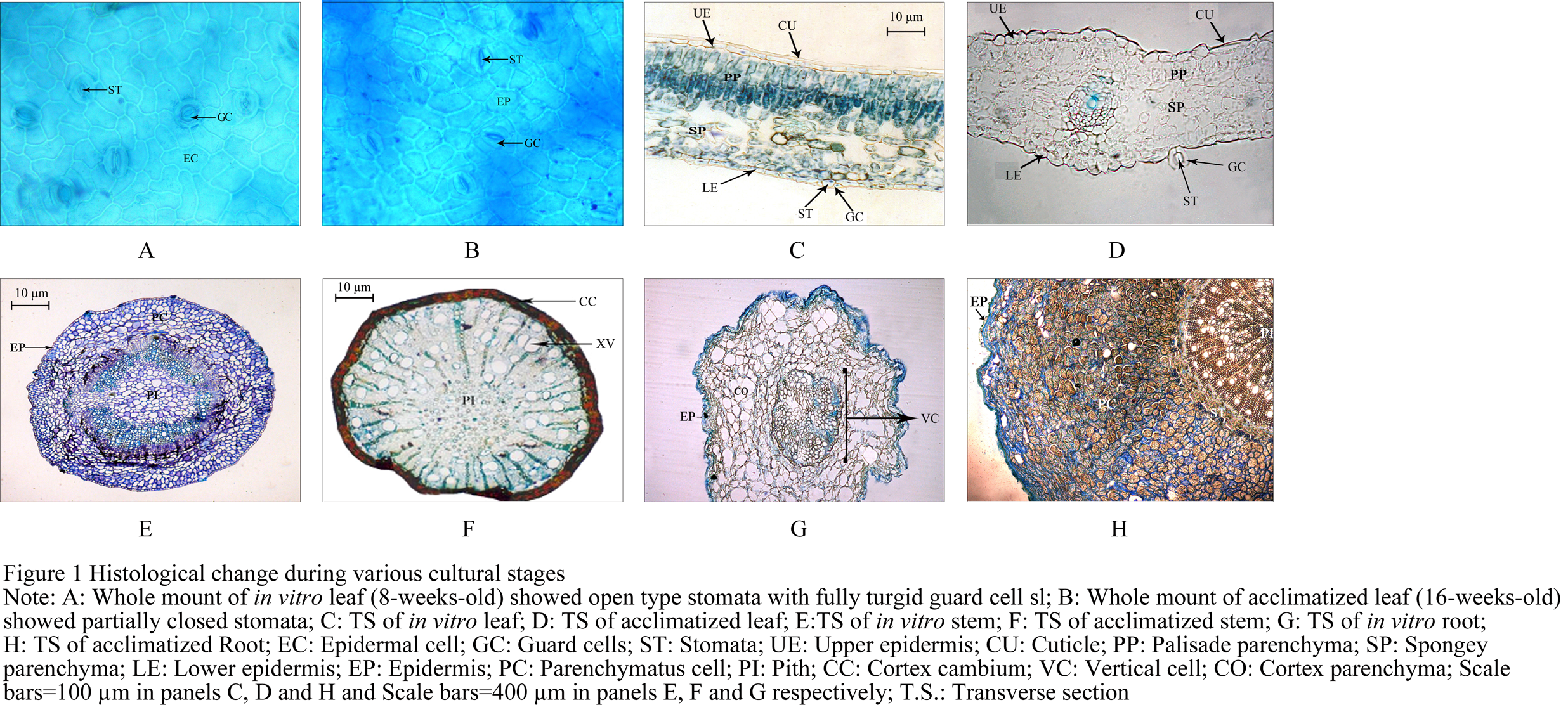
Histology of acclimatized plant leaf (Figure 1B; Figure 1D) showed single layered epidermis with thick cuticle and well developed double layered palisade mesophyll. The lower side had spongy parenchyma with air spaces. Stomata were present in lower epidermis. The in vitro leaf histology showed single layered epidermis with almost no cuticle (Figure 1A; Figure 1C) on both adaxial and abaxial sides. The palisade parenchyma was single layered and poorly developed but the spongy parenchyma was prominent and well developed and the vascular tissue was observed to be poorly developed. It also showed few and open stomatal apparatus with fully turgid guard cells.
|
Figure 1 Histological change during various cultural stages
|
1.3 Histological study of stem
Both the in vitro stem (Figure 1E) and acclimatized stem (Figure 1F) had a polystelic structures. Vascular bundles were arranged in a ring and they were conjoint collateral and open. In the acclimatized stem cork cambium was well developed which was lacking in the in vitro stem. Both the in vitro and acclimatized stems had well developed parenchymatous pith but the pith was mucilaginous in the case of acclimatized stem. The acclimatized stem showed distinct secondary growth with well-developed wood and large woody vessels, while the in vitro stem showed primary medullary rays. The epidermis was uniseriate in both the cases, but it was covered with cuticle in the acclimatized stem, while no cuticle was present in the in vitro stem. The acclimatized stem showed distinct endoderm which was absent in the in vitro stem. The cortex was parenchymatous in the in vitro stem while it was collenchymatous in case of acclimatized Stem.
1.4 Histological study of root
The acclimatized plants root (Figure 1H) had a well-developed parenchymatous cortex which had tannin cells. Pericycle is made of stone cells and there was a prominent casparian strip. The vascular cylinder consisted of secondary xylem towards inner side and secondary phloem towards outer side. The secondary xylem showed wide vessels scattered among trachieds. Medullary rays were present while pith was negligible.
While the in vitro root (Figure 1G) showed poorly developed and undifferentiated cortex with very less amount of storage tissue and no secondary growth at all, the pith was very poorly developed.
Lowest sample area was observed in in vitro stem (10.32 mm2) while highest (19.81 mm2) in acclimatized root. Total thickness of the sample was ranged from (40.4~4.56 µm) with total thickness of the cell v/s parenchymatus cells is also ranges from 1.56~1.97 (Table 2).
| Table 2 Anatomical features of leaf, stem and root of in vitro and acclimatized plantlets |
2 Discussion
2.1 Biochemical analysis
During in vitro phase, it is generally seen that the protein production is low because regenerated plants were photosynthetically inactive therefore protein biosynthesis requirement remains high (Preece and Sutter, 1991; Seon et al., 2000). Lower chlorophyll levels of in vitro plants clearly suggest lower or negligible rate of photosynthesis due to heterotrophic mode of nutrition that ultimately limits the biosynthesis of protein.
During acclimatization in vitro plantlets encounters severe stress condition. The stress response is exhibited in the form of culture reduction and production of heat shock proteins or stress proteins. A decrease in soluble protein content with increasing temperature has been observed in other species (Zhou et al., 1995). Therefore it may be concluded that supplementation of essential nutrients during acclimatization and photoautotrophic development of micropropagated plant has increased survival rate. In vitro plants also showed higher levels of reducing sugars because during micropropagation carbohydrate reserves are built up in leaves by manipulating sucrose in shoot multiplication stage prior to acclimatization. NR is one of the most important metalloflavoprotein regulatory enzyme associated with the process of nitrate assimilation and plant growth which was observed with highest activity in vitro phase, which significantly decreased during acclimatization. Since, NR is substrate inducible enzyme (Haynes, 1964); the presence of high amount nitrate substrate in tissue culture medium probably augments the NR activity during in vitro phase.
2.2 Histological studies
Histological studies of in vitro grown plantlets showed that leaves had more frequent, smaller elongated stomata, more irregular shaped epidermal cells and thin as well as poor developed spongy and palisade parenchyma then the ex vitro plantlets (Yand and Yeh, 2008). Moreover, the leaf surfaces of micropropagated plants were more wettable, suggesting less epicuticular wax deposition on them. These findings suggest that the possible factors those influencing the hardening of plants during the last phases of micropropagation (Gribaudo et al., 2003; Hazel and Harry, 1982). Generally in vitro conditions cause rapid growth and shoot proliferation, thus the plantlet developed shows certain abnormal histological features like altered leaf mesophyll, absence of thick cuticle, sunken or functioning stomata and poor water transport (Majada et al., 1998). Lack of prominent cuticle is a regular feature of in vitro plants and this was proved by the investigation. This cuticle and epicuticle are primary centers for containing water permeability (Pospisilova et al., 2000); this is therefore a major feature leading to excessive water loss and poor survival rate upon transpiration. Our findings are in accordance to earlier reports of Kortemios and Artemios (1997) where in they have reported that Mg2+ and sucrose in the medium influenced the relative size of mesophyll cells, number of chlorophyll and presence of starch grains. The addition of sucrose to the substrate media negatively affected the size of the mesophyll cells under normal Mg2+ concentration (35 mg/L), and positively under high Mg2+ concentration (105 mg/L). Sucrose further resulted in an increase in the numbers of chloroplasts and contained starch grains.
Earlier, Rotondi and Predieri (1996) reported that highest chlorophyll ‘a’ and ‘b’ contents were recorded in leaves of pear cultivars viz. Abbe Fetel and Passe Crassane compact types which also exhibited higher photosynthetic efficiency. During acclimatization transitional forms of leaves, petioles, stems and roots develop that ranged histologically from culture-to control-type. This trend is analogous to the normal developmental sequence of organ formation as it affects the potential for development of successively formed organs (Danielle et al., 1985). Notable changes in leaf histology of bael was observed during different in vitro stages as well as after transferring plantlets to soil. The leaf size increased gradually during micropropagation stages. Similar observations were reported by Brutti et al. (2002) and Apostolo et al. (2005) in Cynara scolymus L. leaves. The in vitro raised plantlets shows poorly developed root histology with no cork cambium, but a single layered epidermis with cortex around the vascular cylinder and vascular bundles were very poorly developed (Petrus-vancea and Sipos, 2010). Non-mucilaginous pith was more prominent, but the xylem parenchyma and fibers were less, making the in vitro raised plantlets more fragile than the acclimatized plantlets. These histological and biochemical observations in in vitro plantlets on comparison with ex vitro plantlets suggest that some procedural changes are necessary for acclimatization of these plants. The successful acclimatization of plantlets may require the full development of leaves, which may largely depend on the rooting stage. This means that in vitro hardening is important and in the present study, we had attempted to select better-suited plants for micropropagation. The supplementation of essential nutrients (½ strength MS plant salt mixture) during acclimatization resulted in increases survival rate proving that photoautotrophic phenomenon has substantial influence on the physiology and development in in vitro regenerated bael plantlets.
3 Materials and methods
3.1 Explant and growth conditions
Three centimeter long nodal stem segments excised from mature tree having at least one axillary bud was used for in vitro regeneration of Aegle marmelos Corr. The nodal explants were washed thoroughly with 2~3 drops of Tween-20 (a liquid detergent) and were rinsed in running tap water for 30 minutes. Explants were treated with 5.24 µM carbendazime + 303.5 µM rifampicin in 100 mL of distilled water and 2~3 drop of Tween-20 for 1 hour in order to disinfect. To overcome the problem of in vitro oxidative browning, the explants were given a pre-treatment with an antioxidant solution comprising ascorbic acid (566 µM), Polyviny lpyrollidine (250×10-2 µM) and citric acid (474.8 µM) for 1 hour. Micropropagation protocol of Pati et al. (2008a; 2008b) was followed to regenerate plantlets of Aegle marmelos Corr.
Eight-week-old rooted plantlets were removed from agar and thoroughly washed using distilled water and transferred to culture bottles (450 mL volume) containing coconut husk. supplemented with ½ strength MS plant salt mixture (Hi-Media, India). Initially the culture bottles were incubated in the culture room under at 4000 lux photo intensity maintained at 25 2℃ temperature for about 15 days, and then shifted to plant growth chamber where, illumination was increased to 10 000 lux with high humidity (70%~80%). The plantlets were then shifted to shade net house (50% shade) provided with misting. The caps of the bottles were loosened gradually and the plants were shifted to pots. Once the plants developed 4~6 leaves, they were shifted to field during the month of July and August.
3.2 Biochemical Studies
The regenerated plantlets of Aegle marmelos Corr were analyzed for different biochemical parameters viz. chlorophyll, reducing sugar, nitrate reductase activity and protein at various regeneration stages (in vitro, acclimatized, field established and control). 500 mg leaf samples were collected from each regeneration stages and used for the biochemical study. The chlorophyll (a, b and total) was estimated as per the method described by Arnon (1949). The reducing sugar was estimated by the method suggested by Ranganna (1986). Nitrate reductase (NR) activity was measured by the method of Srivastava (1975). The total soluble protein was estimated as per the method suggested by Lowery et al. (1951). For all biochemical studis observence were measured by DyNA Quant 200 UV-visible spectrophotometer (Chemito UV-2010, Pvt. Ltd.).
3.3 Histological examination
For histological studies, 0.5 cm2 leaf, stem and root samples of were collected from various cultural stages (8 weeks old in vitro and 16 weeks old acclimatized plants) and fixed into FAA [Formaldehyde (5 mL): Acetic Acid (5 mL): ethanol (90 mL)]. The fixed samples were washed thrice with sterile double distilled water followed by blotted dry on sterile tissue paper. Then fixed samples were dehydrated through the series of t-butyl alcohol in ascending order (30% to 100%). The samples was passed through the gradated ethanol:xylene mixtures (100% t-butyl alcohol, 3:1, 1:1, 1:3 and 100% xylene) using automatic tissue processor electra (Yarco-YSI 104) and embedded in molten paraffin wax ( melting point 56℃) for 8 hrs in order to completely replace the xylene with paraffin wax. 15 m thin sections were cut using microtome (Microm, HM 350S) and stained in 0.1% aqueous toluedine blue O as described by Jensen (1962). The slides were observed under stereoscopic microscope (Leica DFC 320, Japan).
Acknowledgement
We are grateful to Director, CISH, Lucknow for providing facilities.
References
Apostolo N.M., Brutti C.B., and Llorenet B.E., 2005, Leaf anatomy of Cynara scolymus L. in successive micropropagation stages, In vitro Cell Dev. Biol. Plant, 41:307-313
http://dx.doi.org/10.1079/IVP2004606
Arnon D.I., 1949, Copper enzyme in isolated chlorophast, Plant Physiol., 24:1-15
http://dx.doi.org/10.1104/pp.24.1.1 PMid:16654194 PMCid:437905
Brainerd K.E., and Fuchigami L.H., 1982, Stomatal functioning of in vitro and greenhouse aple leaves in darkness, mannitol, ABA and CO2, J. Exp. Bot., 33:388-392
http://dx.doi.org/10.1093/jxb/33.3.388
Brutti C.B., Rubio E.J., Llorente B.E., and Apostolo N.M., 2002, Artichoke leaf morphology and surface features in different micropropagation stages, Biol. Plant, 45:197-204
http://dx.doi.org/10.1023/A:1015132303698
Danielle J., Donnelly, W., Vidaver E. and Kwai Y.L., 1985, The anatomy of tissue cultured red raspberry prior to and after transfer to soil, Plant Cell Tiss. Org. Cult., 34(1):43-50
Gribaudo I., Vallania R., and Novello V., 2003, Anatomical and histological characteristics of the leaves of Vitis vinifera cv. Nebbiolo plants cultivated in vitro and ex vitro, Rivista-di-Frutticoltura-e-di-Ortofloricoltura, 65(2):65-66
Haynes R.J., 1964, Uptake and assimilation of mineral nitrogen by plant. In: Haynes R.J., Ed., Mineral Nitrogen in plant soil system, Academic Press, London. pp.303-378
Hazarika B.N., Nagaraju V., and Parthasarathy V.A, 1996, Morphgenetic response of bale (Aegle marmelos L.) microshoots to benzyl amino purine, Ann Plant Physiol., 10(1): 40
Hazel Y., Wetzstein H., and Sommer E., 1982, Leaf anatomy of tissue cultured Liquidambar styraciflua (Hamamelidaceae) during acclimatization. American J. Bot., 69 (10):1579-1586
http://dx.doi.org/10.2307/2442913
Jensen W.A., 1962, Botanical histochemistry-Principals and practices, W. H. Freeman and Co. San Fransisco and London
Kortessa D.T., and Artemios M.B., 1997, Effects of light, magnesium and sucrose on leaf anatomy, photosynthesis, starch and total sugar accumulation, in kiwifruit cultured in vitro Plant Cell Tiss. Org. Cult., 47:127-134
http://dx.doi.org/10.1007/BF02318948
Lowery O.N., Rosebrough N.J., Farr A.L., and Randall R.J., 1951, Protein measurement with folin phenol reagent, J. Biol. Chem.,193:265-275
Majada J.P., Centeno M.L., Feito I., Fernandez B., and Sanchez-Tames R., 1998, Stomatal and cuticulartraits on carnation tissue cultureunder different ventilation condition, Plant Growth Reg., 25:113-121
http://dx.doi.org/10.1023/B:GROW.0000009706.70078.1c
Maren J.T., and Juniper BE, 1970, The cuticle of plants, St. martin's Press, New York
Mishra M., Pati R., Chandra R., 2006, Clonal micropropagation of Indian gooseberry (Emblica officinalis Gaertn), Ind. J. Genet. Plant Breed, 66(4):361-361
Mishra M., Chandra R., Pati R., and Bajpai A., 2007. Micropropagation of guava (Psidium guajava L.). ISHS Acta Hort, 735:735: 155-158
Pati R., 2008, In vitro plant regeneration and genetic assessment among regenerates using molecular markers in bael (Aegle marmelos Corr.), PhD. Thesis, Awadhesh Pratap Singh University Rewa (M.P.) India
Pati R., Chandra R., Chauhan U.K., Mishra M., and Srivastiva N., 2008a, In vitro clonal propagation of bael (Aegle marmelos Corr.) cv. CISHB1 through enhanced axillary branching. Physiol. Mol. Biol. of Plants, 14(4):337-346
http://dx.doi.org/10.1007/s12298-008-0032-0 PMid:23572900 PMCid:3550643
Pati R., Chandra R., Chauhan U.K., and Mishra M., 2008b, In vitro plant regeneration from mature explant of Aegle marmelos Corr., CV. CISH-B2. Sci. Cult, 74(9-10): 359-367
Pati R., Muthukumar M., 2013, Genetic transformation of bael (Aegle marmelos Corr.), In: Jain S.M., and Gupta D., eds., Biotechnology of neglected and underutilized crops â…§, Springer. [In press: due date April 2013 (ISBN: 978-94-007-5500-0)] http://dx.doi.org/10.1007/978-94-007-5500-0_14
Petrus-vancea A., and Sipos A.M., 2010, Anatomical structure of African violet (Saintpaulia ionantha L.), Analele Universitatii din Oradea-Fascicula Biologie, XVII(2): 219-224
Pospisilova J., Haisel D., Synkova H., Catsky J., Wilhelmova N., Sarka P., Prochazkova D., and Sramek F., 2000, Photosynthetic pigment and gass exchange during ex vitro acclimation of tobacco plants as affected by CO2 supply and abascisic acid, Plant Cell Tiss. Org. Cult., 61:125-133
http://dx.doi.org/10.1023/A:1006402719458
Preece J.C., and Sutter E.G., 1991, Acclimatization of micropropagated plants to greenhouse and field, In: Debergh P.C., and Zimmerman R.H., eds., Micropropagation Technology and Application, Kluwer Academic Pub., The Netherland, pp. 71-93
http://dx.doi.org/10.1007/978-94-009-2075-0_5
Rangana S., 1986, Handbook of analysis and quality control of fruits and vegetable products, New Delhi: Tata Mc-Graw Hill Pub. Co., Ltd., pp.21
Rotondi A., and Predieri S., 1996, Leaf anatomy and photosynthesis of pear trees with different growth habit, Acta Hort., 596
Seon J.H., Cui Y.Y., Kozai T., and Paek K.Y., 2000, Influence of in vitro growth conditions on photosynthetic competence and survival rate of Rehmania glutinosa plantlets during acclimatization period, Plant Cell Tiss. Org. Cult, 61:135-142,
http://dx.doi.org/10.1023/A:1006473223286
Srivastava H.S., 1975, Distribution of nitrate reductase in ageing bean seedling, Plant physiol., 26:995-999
Ticha I., and Radociiova B., 1999, Stomatal morphology during acclimatization of tobacco plantlets to ex vitro conditioned, Biol. Plant, 42:469-474
http://dx.doi.org/10.1023/A:1002118331421 http://dx.doi.org/10.1023/A:1002450210939 http://dx.doi.org/10.1023/A:1002197626878 http://dx.doi.org/10.1023/A:1002166214583
Yand S.H., and Yeh D.M., 2008, In vitro leaf histology, ex virto photosynthetic behaviours and growth of Calathea orbifolia (Linden) Kennedy plants obtained from semisolid medium and temporary immersson systems, Plant Cell. Tiss. Org. Cult., 93:201-207
http://dx.doi.org/10.1007/s11240-008-9363-3
Zhou R.G., Fan Z.H., Li X.Z., Wang Z.W., and Han w., 1995, The effect of heat acclimation on membrane thermostabilityand relative enzyme activity, Acta. Agron. Sinica., 21:568-572
Ziv M., 1986, In vitro hardening and acclimatization of tissue culture plants, In: Withers L.A. and Alderson PG., eds. , Plant tissue culture and its Agricultural application, Butterworths, London, pp.187-196



. PDF(1824KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Rajesh Pati
. Maneesh Mishra
. Ramesh Chandra
. Muthukumar M.
Related articles
. Histology
. Parenchymous cells
. Chlorophyll
. Micropropagation and stomata
Tools
. Email to a friend
. Post a comment